| The evolution of vertebrates shows a trend called cephalization in which special sensory organs develop in the heads of animals, along with the corresponding development of the brain. These special sensory systems, which include the visual, auditory, vestibular, olfactory, and gustatory systems, detect and analyze light, sound, and chemical signals in the environment, as well as signal the position and movement of the head. The stimuli detected and transduced by these systems are most familiar to us when they provide conscious awareness of our environment, but they are equally important as sensory input for reflexive and subconscious behavior.
|
| Vision is one of the most important special senses in humans and, along with audition, is the basis for most human communication. The visual system detects and interprets electromagnetic waves between 400 and 750 nm long, which constitutes visible light.
|
| The eye can distinguish two aspects of light, its brightness (or luminance) and its wavelength (or color). Light enters the eye and impinges on photoreceptors in a specialized sensory epithelium, the retina. The photoreceptors include rods and cones. Rods have high sensitivity for detecting low light intensities but do not provide well-defined visual images, nor do they contribute to color vision. Rods operate best under conditions of reduced lighting (scotopic vision). Cones, by contrast, are not as sensitive to light as rods are and thus operate best under daylight conditions (photopic vision). Cones are responsible for high visual acuity and color vision.
|
| Information processing within the retina is performed by retinal interneurons, and the output signals are carried to the brain by the axons of retinal ganglion cells. The axons travel in the optic nerves; there is a partial crossing in the optic chiasm that results in all input from one side of the visual space being directed to the opposite side of the brain. Posterior to the optic chiasm, the axons of retinal ganglion cells pass through the optic tracts and synapse in nuclei of the brain. The main visual pathway in humans is through the lateral geniculate nucleus (LGN) of the thalamus. This nucleus projects through the visual radiation to the visual cortex. Other visual pathways project to the superior colliculus, pretectum, and hypothalamus, and these structures participate in orientation of the eyes, control of pupil size, and circadian rhythms, respectively.
|
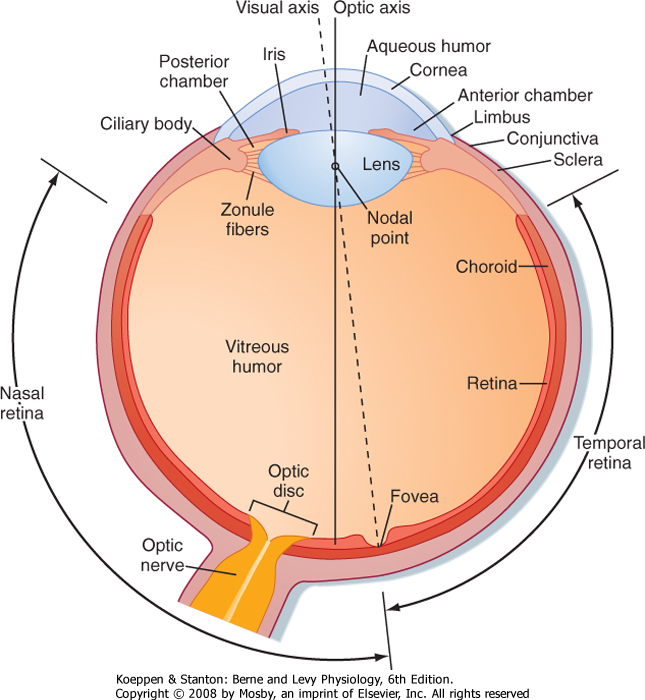
| The wall of the eye is composed of three concentric layers (Fig. 8-1). The outer layer, or the fibrous coat, includes the transparent cornea, with its epithelium (the conjunctiva), and the opaque sclera. The middle layer, or vascular coat, includes the iris and the choroid. The iris contains both radially and circularly oriented smooth muscle fibers, which make up the pupillary dilator and sphincter muscles. The choroid is rich in blood vessels that support the outer layers of the retina, and it also contains pigment. The innermost layer of the eye is the retina, which is embryologically derived from the diencephalon and is therefore part of the central nervous system (CNS). The functional part of the retina covers the entire posterior aspect of the eye except for the optic nerve head or optic disc, which is where the optic nerve axons leave the retina. Because there are no receptors at this location, it is often referred to as the anatomic "blind spot" (Fig. 8-1).
|
| A number of functions of the eyes are under muscular control. Externally attached extraocular muscles aim the eyes toward an appropriate visual target (see Chapter 9). These muscles are innervated by the oculomotor (cranial nerve [CN] III), trochlear (CN IV), and abducens (CN VI) nerves. Several muscles are also found within the eye (intraocular muscles). The muscles in the ciliary body control lens shape and thereby the focus of images on the retina. The pupillary dilator and sphincter muscles allow the iris to control the amount of light entering the eye, similar to the diaphragm of a camera. The dilator is activated by the sympathetic nervous system, whereas the sphincter and ciliary muscles are controlled by the parasympathetic nervous system (through the oculomotor nerve) (see Chapter 11).
|
| page 123 |  | | page 124 |
| Figure 8-1 View of a horizontal section of the right eye. (Redrawn from Wall GL: The Vertebrate Eye and Its Adaptive Radiation. Bloomfield Hills, MI, Cranbrook Institute of Science, 1942.) |
| If aqueous humor is not absorbed adequately, intraocular pressure increases, a condition known as glaucoma. An increase in intraocular pressure can cause blindness by impeding blood flow to the retina. In addition, cloudiness or objects floating (floaters or "mouches volantes") in the vitreous humor can disrupt the light path to the retina and distort clear vision. |
|
| Light enters the eye through the cornea and passes through a series of transparent fluids and structures that are collectively called the dioptric media. These fluids and structures consist of the cornea, aqueous humor, lens, and vitreous humor. The aqueous humor, located in the anterior and posterior chambers and the vitreous humor in the space behind the lens,
respectively, help maintain the shape of the eye. The aqueous humor is secreted by the epithelium of the ciliary body into the posterior chamber of the eye. It then circulates through the pupil and into the anterior chamber, where it is drained into the venous system by the canal of Schlemm. Aqueous humor pressure, which is normally less than 22 mm Hg, determines the pressure within the eye. The vitreous humor is a gel composed of extracellular fluid that contains collagen and hyaluronic acid; unlike aqueous humor, however, it turns over very slowly.
|
| Normally, light from a visual target is focused sharply on the retina by the cornea and lens, which bend or refract the light. The cornea is the major refractive element of the eye, with a refractive power of 43 diopters* (D). However, unlike the cornea, the lens can change shape and vary its refractive power between 13 and 26 D. Thus, the lens is responsible for adjusting the optical focus of the eye. Suspensory ligaments (or zonule fibers), which attach to the wall of the eye at the ciliary body (Fig. 8-1), hold the lens in place. When the muscles in the ciliary body are relaxed, the tension exerted by the suspensory ligaments flattens the lens. When the ciliary muscles contract, the tension on the suspensory ligaments is reduced; this process allows the somewhat elastic lens to assume a more spherical shape. The ciliary muscles are activated by the parasympathetic nervous system (via the oculomotor nerve).
|
| In this way the lens allows the eye to focus on, or accommodate to, either near or distant objects. For instance, when light from a distant visual target enters a normal eye (one with a relaxed ciliary muscle), the target is in focus on the retina. However, if the eye is directed at a nearby visual target, the light is initially focused behind the retina (i.e., the image at the retina is blurred) until accommodation occurs. The ciliary muscle contracts and the zonule fibers relax; the image is sharpened when the convexity of the lens increases as a result of these muscular changes.
|
| page 124 | | | page 125 |
| Although the optic axis of the human eye passes through the nodal point of the lens and reaches the retina at a point between the fovea and the optic disc (Fig. 8-1), the eye is directed by the oculomotor system to a point, called the fixation point, on the visual target. Light from the fixation point passes along the optic axis, through the nodal point, and is focused on
the fovea. Light from the remainder of the visual target falls on the retina surrounding the fovea.
|
| As an individual ages, the elasticity of the lens gradually declines. As a result, accommodation of the lens for near vision becomes progressively less effective, a condition called presbyopia. A young person can change the power of the lens by as much as 14 D. However, by the time that a person reaches 40 years of age, the amount of accommodation halves, and after 50 years it decreases to 2 D or less. Presbyopia can be corrected by convex lenses. |
| Defects in focus are caused by a discrepancy between the size of the eye and the refractive power of the dioptric media. For example, in myopia (near-sightedness), the images of distant objects are focused in front of the retina. Concave lenses correct this problem. Conversely, in hypermetropia (far-sightedness), the images of distant objects are focused behind the retina; this problem can be corrected with convex lenses. In astigmatism, an asymmetry exists in the radii of curvature of different meridians of the cornea or lens (or sometimes of the retina). Astigmatism can often be corrected with lenses that possess appropriately matched radii of curvature. |
|
| Proper focus of light on the retina depends not only on the lens but also on the iris, which also adjusts the amount of light that can enter the eye. In this respect
the iris acts like the diaphragm in a camera, which also controls the depth of field of the image and the amount of spherical aberration produced by the lens. When the pupil is constricted, the depth of field is increased, and the light is directed through the central part of the lens, where spherical aberration is minimal. Pupillary constriction occurs reflexively when the eye accommodates for near vision or adapts to bright light, or both. Thus, when a person reads or does other fine visual work, the quality of the image is improved by having adequate light.
|
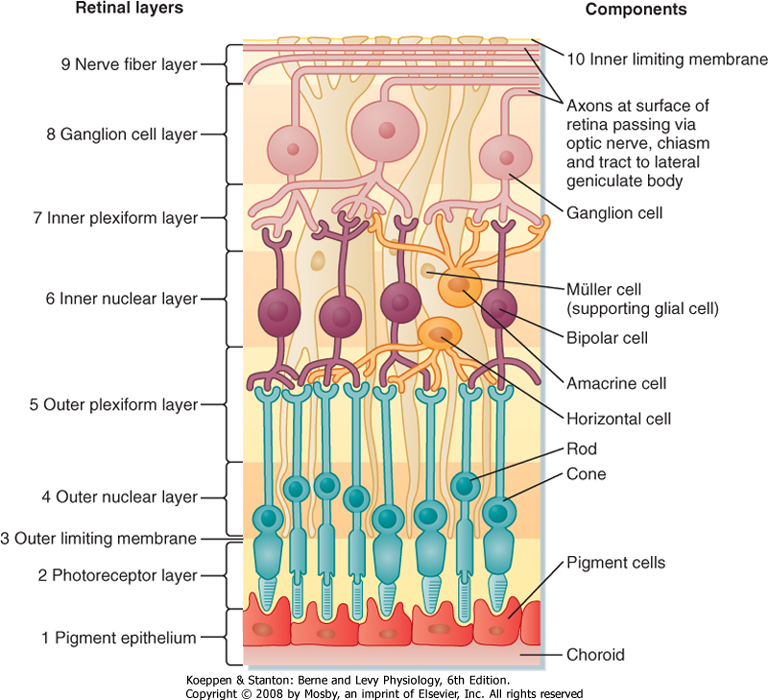
| The 10 layers of the retina are shown in Figure 8-2. The retina begins with the pigmented epithelium (layer 1), which is just inside the choroid. The pigment cells have tentacle-like processes that extend into the photoreceptor layer (layer 2) and surround the outer segments of the rods and cones. These processes prevent transverse scatter of light between photoreceptors. In addition, they serve a mechanical function in maintaining contact between layers 1 and 2 so that the pigmented epithelium can (1) provide nutrients and remove waste from the photoreceptors; (2) phagocytose the ends of the outer segments of the rods, which are continuously shed; and (3) reconvert metabolized photopigment into a form that can be reused after it is transported back to the photoreceptors.
|
| Figure 8-2 Layers of the retina. Light impinging on the retina comes from the top of the figure and passes through all the superficial layers to reach the photoreceptor rods and cones. |
| page 125 | | | page 126 |
| The junction between layers 1 and 2 of the retina in adults represents the surface of contact between the anterior and posterior walls of the embryonic optic cup during development and is structurally weak. Retinal detachment is separation at this surface and can cause loss of vision because of displacement of the retina from the focal plane of the eye. It can also lead to the death of photoreceptor cells, which are maintained by the blood supply of the choroid (the photoreceptor layer itself is avascular). Deterioration of the pigmented epithelium can also result in macular degeneration, a critical loss of high-acuity central and color vision without affecting peripheral vision. |
|
| Light rays that originate from different parts of the visual target map onto the photoreceptor array of
layer 2 in a point-to-point fashion. Retinal glial cells, known as Müller cells, play an important role in maintaining the internal geometry of the retina. Müller cells are oriented radially, parallel to the light path through the retina. The outer ends of Müller cells form tight junctions with the inner segments of the photoreceptors. The numerous connections made between Müller cells and the inner segments give the appearance of a continuous layer, the outer limiting membrane (layer 3 of the retina).
|
| Inside the external limiting membrane is a layer of nuclei called the outer nuclear layer (layer 4 of the retina) that contains the cell bodies and nuclei of the rods and cones. The next layer of the retina (layer 5) is called the outer plexiform layer. It contains synapses between the photoreceptors and retinal interneurons, including the bipolar cells and horizontal cells, whose cell bodies are found in the inner nuclear layer (layer 6 of the retina). This layer also contains the cell bodies of other retinal interneurons (the amacrine and interplexiform cells) and the Müller cells.
|
| The next layer is the inner plexiform layer (layer 7 of the retina). It contains synapses between the retinal neurons of the inner nuclear layer, including the bipolar and amacrine cells, and the ganglion cells. Layer 8 of the retina is the ganglion cell layer. As previously mentioned, the ganglion cells are the output cells of the retina; it is their axons that transmit visual information to the brain. These axons form the optic fiber layer (layer 9 of the retina), pass along the vitreous surface of the retina while avoiding the fovea, and enter the optic disc, where they leave the eye in the optic nerve. The portions of the ganglion cell axons that are in the optic fiber layer remain unmyelinated, but the axons become myelinated after they reach the optic disc. The lack of myelin where the axons cross the retina is a specialization that helps permit light to pass through the inner retina with minimal distortion.
|
| The innermost layer of the retina is the inner limiting membrane (layer 10 of the retina). This layer is formed by the end-feet of Müller cells.
|
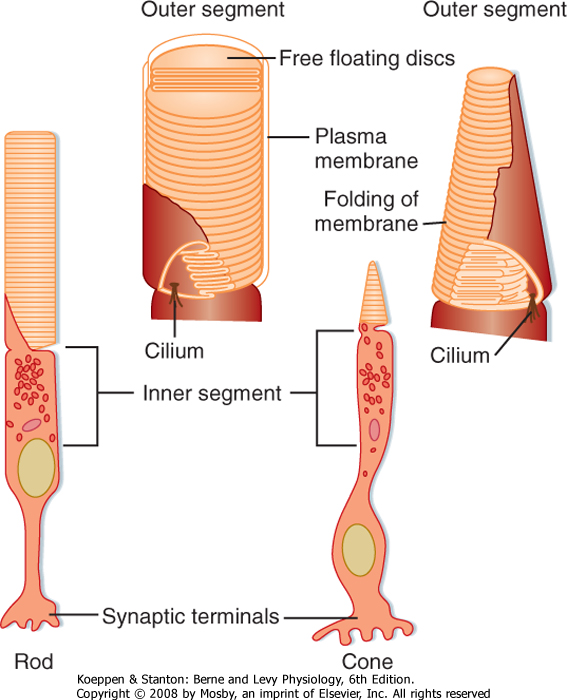
| Figure 8-3 Rods and cones. The drawings at the bottom show the general features of a rod and a cone. The insets show the outer segments. |
| Structure of Photoreceptors: Rods and Cones
|
| Each rod or cone photoreceptor cell is composed of a cell body (in layer 4), an inner segment and an outer segment that extend into layer 2, and a synaptic terminal that projects into layer 5 (Fig. 8-3). The outer segments of cones are not as long as those of rods, and they contain stacks of disc membranes formed by infoldings of the surface membrane. The outer segments of rods are longer, and the stacks of membrane discs float freely in the outer segment after having disconnected from the surface membrane when formed at the base. Both sets of discs are rich in photopigment molecules, but the greater photopigment density of rods accounts for their greater sensitivity to light. A single photon can elicit a rod response, whereas several hundred photons may be required for a cone response.
|
| The inner segments of the photoreceptors are connected to the outer segments by a modified cilium that contains nine pairs of microtubules, but it lacks the two central pairs of microtubules found in most cilia. The inner segments contain a number of organelles, including numerous mitochondria.
|
| The photopigment is synthesized in the inner segment and incorporated into the membranes of the outer segment. In rods, the pigment is inserted into new membranous discs, which are then displaced distally until they are eventually shed at the apex of the outer segment. There, they are phagocytozed by cells of the pigmented epithelium. This process determines the rod-like shape of the outer segments of rods. In cones, the photopigment is inserted randomly into the membranous folds of the outer segment, and shedding, comparable to that seen in rods, does not take place.
|
| page 126 | | | page 127 |
| Regional Variations in the Retina
|
| The macula lutea is the area of central vision and is characterized by a slight thickening and a pale color. The thickness is due to the high concentration of photoreceptors and interneurons, which are needed for high-resolution vision. The pale color is a consequence of the fact that both optic nerve fibers and blood vessels are routed around it.
|
| The fovea, which is a depression in the macula lutea, is the region of the retina that has the very highest visual resolution. Correspondingly, the image from the fixation point is focused on the fovea. The retinal layers in the foveal region are unusual because several of them appear to be pushed aside into the surrounding macula. Because light can reach the foveal photoreceptors without having to pass through the inner layers of the retina, both image distortion and light loss are minimized. The fovea has cones with unusually long and thin outer segments. This cone shape permits high packing density. In fact, cone density is maximal in the fovea, and this high density provides for high visual resolution, as well as high quality of the image (Fig. 8-4).
|
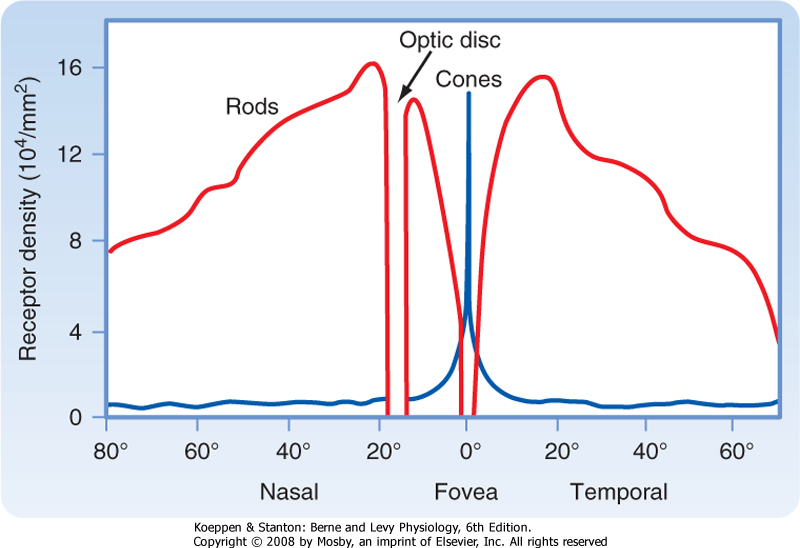
|
| Figure 8-4 This graph plots the density of cones and rods as a function of retinal eccentricity from the fovea. Note that cone density peaks at the fovea, rod density peaks at about 20 degrees eccentricity, and no photoreceptors are found at the optic disc. (Data from Cornsweet TN: Visual Perception. New York, Academic Press, 1970.) |
| The optic disc lacks photoreceptors and therefore lacks photosensitivity. Thus, the optic disc is a "blind spot" in the visual surface of the retina. A person is normally unaware of the blind spot, both because the corresponding part of the visual field can be seen by the contralateral eye and because of the psychological process in which incomplete visual images tend to be completed perceptually.
|
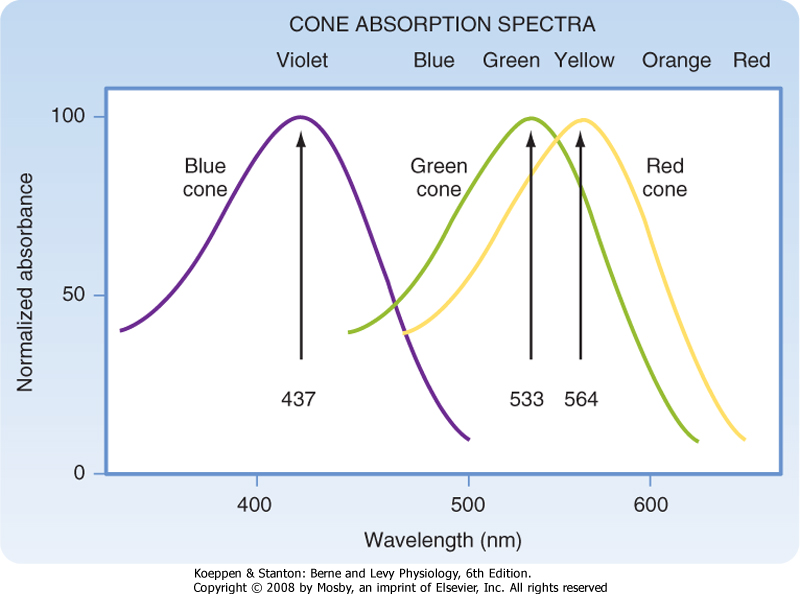
| Light energy must be absorbed for it to be detected by the retina. Light absorption is accomplished by the visual pigments, which are located in the outer segments of the rods and cones. The pigment found in the outer segments of rods is rhodopsin, or visual purple (so named because it has a purple appearance after green or blue light have been absorbed), and it absorbs light best at a wavelength of 500 nm. Three variants of visual pigment are found in cones, and these cone pigments absorb best at 437 nm (blue), 533 nm (green), or 564 nm (red). However, the absorption spectrum of visual pigments is broad so that they overlap considerably (Fig. 8-5).
|
| Figure 8-5 The spectral sensitivity of the three types of cone pigments in the human retina is shown. Note that the curves overlap. (Data from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.) |
| page 127 | | | page 128 |
| Rhodopsin is formed when a retinal isomer, 11-cis retinal, is combined with a glycoprotein known as opsin. When rhodopsin absorbs light, it is "boosted" to a higher energy state. This boost causes a series of chemical changes that lead to isomerization of 11-cis retinal to all-trans retinal, release of the bond with opsin, and conversion of retinal to retinol. Separation of all-trans retinal from opsin causes bleaching of the visual pigment; that is, the pigment loses its purple color.
|
| As mentioned, the axons of retinal ganglion cells cross the retina in the optic fiber layer (layer 9) and enter the optic nerve at the optic disc. Axons in the optic fiber layer pass around the macula and fovea, as do the blood vessels that supply the inner layers of the retina. The optic disc can be visualized on physical examination with an ophthalmoscope. The normal optic disc has a slight depression in its center. Changes in the appearance of the optic disc are important clinically. For example, the depression may be exaggerated by loss of ganglion cell axons (optic atrophy), or the optic disc may protrude into the vitreous space because of edema (papilledema) caused by increased intracranial pressure. |
|
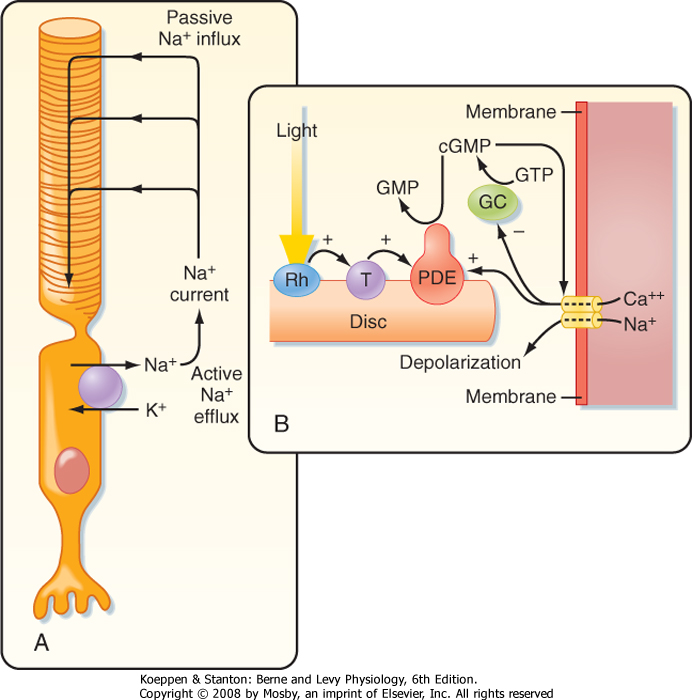
|
| Figure 8-6 A, Drawing of a rod. The flow of current in the dark is indicated, as well as the Na+ pump. B, Sequence of the second messenger events that follow the absorption of light. cGMP, cyclic guanosine monophosphate; GC, guanylate cyclase; GTP, guanosine triphosphate; PDE, phosphodiesterase; Rh, rhodopsin; T, transducin. |
| In darkness, photoreceptors are slightly depolarized (around -40 mV) because cGMP-gated Na+ channels (Fig. 8-6A) in their outer segments are open, thereby increasing gNa and driving the membrane
potential toward the Na+ equilibrium potential. This net influx of Na+ results in a continuous current, called the dark current. As a consequence of this constant depolarization, the neurotransmitter glutamate is tonically released at the rod cell's synapses. Intracellular [Na+] is kept at a steady-state level by the pumping action of Na+,K+-ATPase.
|
| When light is absorbed, the photoisomerization of rhodopsin activates a G protein called transducin (Fig. 8-6B). This G protein, in turn, activates cyclic guanosine monophosphate phosphodiesterase, which is associated with the rhodopsin-containing discs, hydrolyzes cGMP to 5'-GMP, and lowers the cGMP concentration in the rod cytoplasm. The reduction in cGMP leads to closing of the cGMP-gated Na+ channels, hyperpolarization of the photoreceptor membrane, and a reduction in the release of transmitter. Thus, cGMP acts as a "second messenger" to translate reception of a photon by the photopigment into a change in membrane potential.
|
| Rhodopsin contains a chromophore, called retinal, that is the aldehyde of retinol, or vitamin A. Retinol is derived from carotenoids, such as β-carotene, the orange pigment found in carrots. Like other vitamins, retinol cannot be synthesized by humans; instead, it is derived from food sources. Individuals with a severe vitamin A deficiency suffer from "night blindness," a condition in which vision is defective in poor illumination. |
| page 128 | | | page 129 |
| The extraordinary sensitivity of rods, which can signal the capture of a single photon, is enhanced by an amplification mechanism such that photoactivation of only one rhodopsin molecule can activate hundreds of transducin molecules. In addition, each phosphodiesterase molecule hydrolyzes thousands of cGMP molecules per second. Similar events occur in cones, but the membrane hyperpolarization occurs much more quickly than in rods, and requires thousands of protons.
|
| Thus, in all photoreceptors, capture of light energy leads to (1) hyperpolarization of the photoreceptor and (2) a reduction in the release of transmitter. Note that with the very short distance between the site of transduction and the synapse, this transmitter modulation is accomplished without the generation of an action potential.
|
| Adaptation permits the retina to adjust its sensitivity to large changes in ambient lighting, such as you experience when entering a darkened movie theater or, later, leaving to encounter afternoon sunlight. Light adaptation is associated with a reduction in the amount of rhodopsin and the resulting reduced photosensitivity. In bright light, 11-cis retinal is isomerized into the all-trans form, which then splits from the opsin. To regenerate the rhodopsin, the all-trans retinal is transported to the retinal pigmented cell layer to be reduced to retinol, isomerized, and esterified back to 11-cis retinal. It is then transported back to the photoreceptor layer, taken up by outer segments, and recombined with opsin to regenerate the rhodopsin. Light adaptation, which occurs rapidly, within seconds, favors cone vision because the rhodopsin in rods bleaches (separates from its opsin) more readily than the cone pigments do.
|
| The regeneration of photopigment is also involved in dark adaptation, a process that results in an increase in visual sensitivity. Cones adapt more rapidly to darkness than rods do, but their adapted threshold is relatively high. Thus, cones do not function when the ambient light level is low. By contrast, rods adapt to darkness slowly, but their sensitivity increases. Within 10 minutes in a dark room, rod vision is more sensitive than cone vision.
|
| Dark adaptation is very familiar to moviegoers, who must wait several minutes after entering the darkened theater before they can see an empty seat. Although the theater is dark and rod vision is operative, visual acuity is low and colors are not distinguished (this is called scotopic vision). When the movie is projected, however, cone function resumes (this is called photopic vision), and visual acuity and color vision are restored.
|
| The three visual pigments in the cone outer segments have opsins that differ from the opsin found in rhodopsin. As a result of these differences, the three types of cone pigments absorb light best at different wavelengths. Although the cone pigments have maximum efficiency closer to violet, green, and yellow wavelengths, they are referred to as blue, green, and red pigments, respectively (Fig. 8-5).
|
| According to the trichromacy theory, these differences in absorption efficiency are presumed to account for color vision because a suitable mixture of three colors can produce any other color. However, a neural system must also exist for the analysis of color brightness because the amount of light absorbed by a visual pigment, as well as the subsequent response of the cell, depends on both the wavelength and the intensity of the light (Fig. 8-5). Two or three of the cone pigments may absorb a particular wavelength of light, but the amount absorbed by each will differ according to their efficiencies at that wavelength. If the intensity of the light is increased (or decreased), all will absorb more (or less), but the ratio of absorption among them will remain constant. Consequently, there must be a neural mechanism to compare the absorption of light of different wavelengths by the different types of cones for the visual system to distinguish different colors. At least two different kinds of cones are required for color vision. The presence of three kinds decreases the ambiguity in distinguishing colors when all three absorb light, and it ensures that at least two types of cones will absorb most wavelengths of visible light.
|
| The opponent process theory is based on observations that certain pairs of colors seem to activate opposing neural processes. Green and red are opposed, as are yellow and blue, as well as black and white. For example, if a gray area is surrounded by a green ring, the gray area appears to acquire a reddish color. Furthermore, a greenish red or a bluish yellow color does not exist. These observations are supported by findings that neurons activated by green are inhibited by red. Similarly, neurons excited by blue may be inhibited by yellow. Neurons with these characteristics are found both in the retina and at higher levels of the visual pathway and seem to serve to increase our ability to see the contrast between opposing colors.
|
| Observations on color blindness are consistent with the trichromacy theory. In color blindness, a genetic defect (sex-linked recessive), one or more cone mechanisms are lost. Normal people are trichromats because they have three cone mechanisms. Individuals who have lost one of the cone mechanisms are called dichromats. When the long-wavelength cone mechanism is lost, the resulting condition is called protanopia; loss of the medium-wavelength system causes deuteranopia; and loss of the short-wavelength system causes tritanopia. Monochromats have lost all three cone mechanisms (or in some cases, two of them). |
| page 129 | | | page 130 |
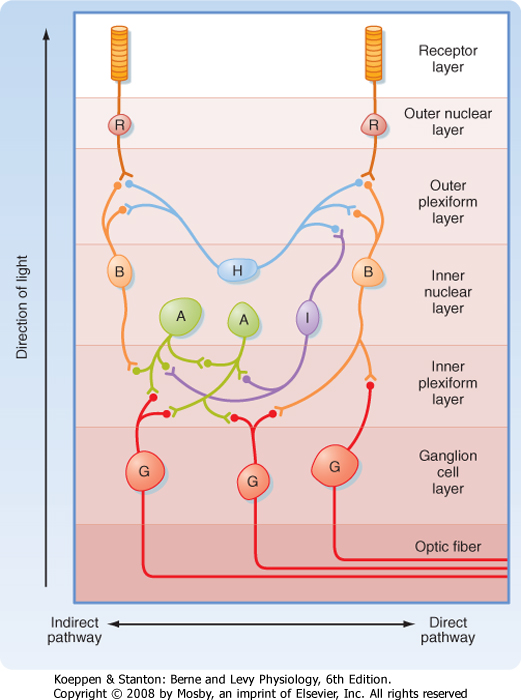
| Figure 8-7 Basic retinal circuitry. The arrow at the left indicates the direction of light through the retina. A, amacrine cells; B, bipolar cells; G, ganglion cells; H, horizontal cells; I, interplexiform cells; R, photoreceptors. |
| A diagram of the basic circuitry of the retina is shown in Figure 8-7. Photoreceptors (R) synapse on the dendrites of bipolar cells (B) and horizontal cells (H) in the outer plexiform layer. The horizontal cells make
reciprocal synaptic connections with bipolar cells, are electrically coupled to other horizontal cells, and receive input from interplexiform cells (I). Bipolar cells synapse on the dendrites of ganglion cells (G) and on the processes of amacrine cells (A) in the inner plexiform layer. Amacrine cells connect with ganglion cells, other amacrine cells, and interplexiform cells.
|
| Several features of this circuitry are noteworthy. Input to the retina is provided by light striking the photoreceptors. The output is carried by axons of the retinal ganglion cells to the brain. Information is processed within the retina by the interneurons. The most direct pathway through the retina is from a photoreceptor to a bipolar cell and then to a ganglion cell (Fig. 8-7). More indirect pathways that provide for intraretinal signal processing involve photoreceptors, bipolar cells, amacrine cells, and ganglion cells, as well as horizontal cells to provide lateral interactions between adjacent pathways. Interplexiform cells allow interactions to occur from the inner to the outer retina.
|
| Contrasts in Rod and Cone Pathway Functions
|
| Rod and cone pathways have several important functional differences, based partly on differences in their phototransduction mechanisms and partly on retinal circuitry. As described previously, rods have more photopigment and a better signal amplification system than cones do, and there are many more rods than cones. Thus, rods function better in dim light (scotopic vision), and loss of rod function results in night blindness. In addition, all rods contain the same photopigment, so they cannot signal color differences. Furthermore, because many rods converge onto individual bipolar cells, thereby resulting in very large receptive fields, rods cannot provide high-resolution vision. Finally, in bright light most rhodopsin is bleached, so rods no longer function under photopic conditions.
|
| page 130 | | | page 131 |
| Cones have a higher threshold to light and thus are not activated in dim light after dark adaptation. However, they operate very well in daylight. They provide high-resolution vision because only a few cones converge onto individual bipolar cells in the cone pathways. Moreover, no convergence occurs in the fovea where the cones make one-to-one connections to bipolar cells. As a result of the reduced convergence, cone pathways have very small receptive fields and can resolve stimuli that originate from sources very close to each other. Cones also respond to sequential stimuli with good temporal resolution. Finally, cones have three different cone photopigments. Thus, they can discriminate relative spectral content independent of absolute intensity and therefore provide for color vision. Loss of cone function
results in functional blindness; rod vision is not sufficient for normal visual requirements.
|
| The distances between retinal components are short. Hence, modulated transmitter release and postsynaptic potentials are sufficient for most of the activity in retinal circuits, and action potentials are not required in most of the interneurons. Only the ganglion cells and some amacrine cells generate action potentials. It is unclear why amacrine cells have action potentials, but ganglion cells must generate them to transmit information over the relatively long distance from the retina to the brain.
|
| Although receptor potentials in photoreceptors are hyperpolarizing, synaptic potentials in the retina can be either hyperpolarizing or depolarizing. Hyperpolarizing events reduce neurotransmitter release from the synaptic terminals of a retinal interneuron, whereas depolarizing events increase neurotransmitter release.
|
| Receptive Field Organization
|
| The receptive field of an individual photoreceptor is small and circular. Light in the receptive field will hyperpolarize the photoreceptor cell and cause it to release less neurotransmitter. The receptive fields of photoreceptors and retinal interneurons determine the receptive fields of the retinal ganglion cells onto which their activity converges. The characteristics of the receptive fields of retinal ganglion cells constitute an important step in visual information processing because it is this processed information about visual events that is conveyed to the brain.
|
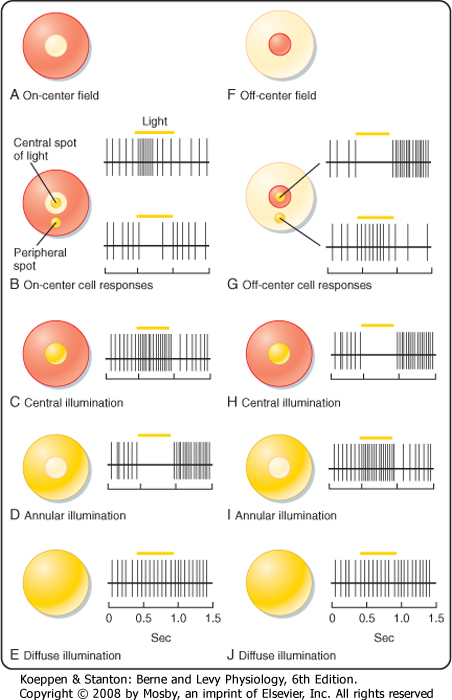
| Figure 8-8 The receptive fields of on-center (A) and off-center (F) bipolar cells and their responses to central (B, C, and G, H), surround (B, D, and G, I), and diffuse (E, J) illumination in their receptive fields. (Redrawn from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.) |
| page 131 | | | page 132 |
| A bipolar cell that receives input from a photoreceptor can have either of two types of receptive fields, as shown in Figure 8-8. Both are described as having a center-surround organization in which the light that
strikes the central region of the receptive field either excites or inhibits the cell, whereas the light that strikes the annular region that surrounds the central portion has the converse effect. The receptive field with a centrally located excitatory region surrounded by an inhibitory annulus is called an on-center, off-surround receptive field (Fig. 8-8, A to E). Bipolar cells with such a receptive field are described as "on" bipolars. The other type of receptive field has an off-center, on-surround arrangement, which characterizes "off" bipolars (Fig. 8-8, F to J).
|
| The receptive fields of bipolar cells depend on input from photoreceptors and from horizontal cells. The neurotransmitter used in the retinal pathway from photoreceptor cells to bipolar cells and to horizontal cells is the excitatory amino acid glutamate. Excitatory amino acids depolarize "off" bipolar cells, as well as horizontal cells, through the activation of ionotropic glutamate receptors. These are called "off" bipolars because when light is removed from the receptive field center, the photoreceptor is depolarized and releases more glutamate to depolarize the bipolar cell. In contrast, "on" bipolar cells have metabotropic glutamate receptors that close their Na+ channels, and thus "on" bipolars are depolarized by turning the light on because the reduced release of glutamate results in more influx of Na+.
|
| In other words, if the neurotransmitter tonically released by the photoreceptor hyperpolarizes the bipolar cell, absorption of light will hyperpolarize the photoreceptor and thereby reduce its release of the neurotransmitter; the "on" bipolar cell will be depolarized (disinhibited) and thus excited. On the other hand, the neurotransmitter tonically released by the photoreceptor depolarizes the "off" bipolar cell, and it will be hyperpolarized (disfacilitated) by central illumination.
|
| The central property of bipolar cell receptive fields is due to only a few directly connected photoreceptors. The antagonistic surround response is due to light impinging on adjacent photoreceptors, which changes the activity of horizontal cells. This pathway through the horizontal cells results in a response that is opposite in sign to that produced directly by the photoreceptors that mediate the center response. The basis for this is that horizontal cells, like "off" bipolars, are hyperpolarized in the light and, because they are electrically coupled to each other by gap junctions, have very large receptive fields. Illumination in the periphery of a bipolar cell's receptive field (such as by an annulus that does not affect the photoreceptor to which it is directly connected) will stimulate neighboring photoreceptors and hyperpolarize the horizontal cells. The hyperpolarized horizontal cells release less glutamate onto bipolars and photoreceptors. This tends to depolarize the photoreceptors and mimics darkness such that "on" bipolars are inhibited and "off" bipolars are excited (Fig. 8-8).
|
| Bipolar cells may not respond at all to large or diffuse areas of illumination, covering both the receptors that cause the surround response and those responsible for the center response because of the opposing actions from the center and surround (Fig. 8-8, E, J). Thus, bipolar cells may not signal changes in the intensity of light that strikes a large area of the retina. On the other hand, a small spot of light moving across the receptive field may sequentially and dramatically alter the activity of the bipolar cell as the light crosses the receptive field from surround to center and then back again to surround. This demonstrates that bipolar cells respond best to the local contrast of stimuli and function as contrast detectors.
|
| Amacrine cells receive input from different combinations of on-center and off-center bipolar cells. Thus, their receptive fields are mixtures of on-center and off-center regions. There are many different types of amacrine cells, and they may use at least eight different neurotransmitters. Accordingly, the contributions of amacrine cells to visual processing are complex.
|
| Ganglion cells may receive dominant input from bipolar cells, dominant input from amacrine cells, or mixed input from amacrine and bipolar cells. When amacrine cell input dominates, the receptive fields of ganglion cells tend to be diffuse, and they are either excitatory or inhibitory. Most ganglion cells are dominated by bipolar cell input and have a center-surround organization, similar to that of bipolar cells.
|
| Experiments have shown that in primates, retinal ganglion cells can be subdivided into three general types called P cells, M cells, and W cells. P and M cells are fairly homogeneous groups, whereas W cells are heterogeneous. P cells are so named because they project to the parvocellular layers of the LGN, whereas M cells project to the magnocellular layers of the LGN. P and M cells have center-surround receptive fields; hence, they are presumably controlled by bipolar cells. W cells may also have center-surround receptive fields, but many have large, diffuse receptive fields (which corresponds to extensive dendritic fields) and slowly conducting axons, and they respond poorly to visual stimuli. They are probably influenced chiefly through amacrine cell pathways, but less is known of them than of M and P cells.
|
|
Table 8-1.
Properties of Retinal Ganglion Cells |
| Properties | P Cells | M Cells | W Cells |
| Cell body and axon | Medium sized | Large | Small |
| Dendritic tree | Restricted | Extensive | Extensive |
| Receptive field |
| Size | Small | Medium | Large |
| Organization | Center-surround | Center-surround | Diffuse Poorly responsive |
| Adaptation | Tonic | Phasic | |
| Linearity | Linear | Nonlinear | |
| Wavelength | Sensitive | Insensitive | Insensitive |
| Luminance | Insensitive | Sensitive | Sensitive |
| page 132 | | | page 133 |
| Several of the physiological differences among these cell types correspond to morphological differences (Table 8-1). For example, P cells have small receptive
fields (which corresponds to smaller dendritic trees) and more slowly conducting axons than M cells do. In addition, P cells show a linear response in their receptive field; that is, they respond with a sustained, tonic discharge of action potentials to maintained light but do not signal shifts in the pattern of illumination as long as the overall level of illumination is constant. Thus, a small object entering a P cell's central receptive field will change its firing, but continued movement within the field will not be signaled. P cells respond differently to different wavelengths of light. Because there are blue, green, and red cones, many combinations of color properties are possible, but in fact P cells have been shown to have only opposing responses to red and green or to blue and yellow (a combination of red and green). They may have center-surround antagonism in which one color excites the center while the other inhibits the surround (or vice versa), or one color might excite the entire receptive field while another inhibits it (e.g., R+G- describes a cell that is excited by red and inhibited by green). These mechanisms can greatly reduce the ambiguity of color detection caused by the overlap in cone color sensitivity and may provide a substrate for the opponency process observations.
|
| M cells, on the other hand, respond with phasic bursts of action potentials to the redistribution of light, such as would be caused by the movement of an object within their large receptive fields. M cells are not sensitive to differences in wavelength but are more sensitive to luminance than P cells are.
|
| Thus, the output of the retina consists primarily of ganglion cell axons from (1) sustained, linear P cells with small receptive fields that convey information about color, form, and fine details and (2) phasic, nonlinear M cells with larger receptive fields that convey information about illumination and movement. Both come in on-center and off-center varieties.
|
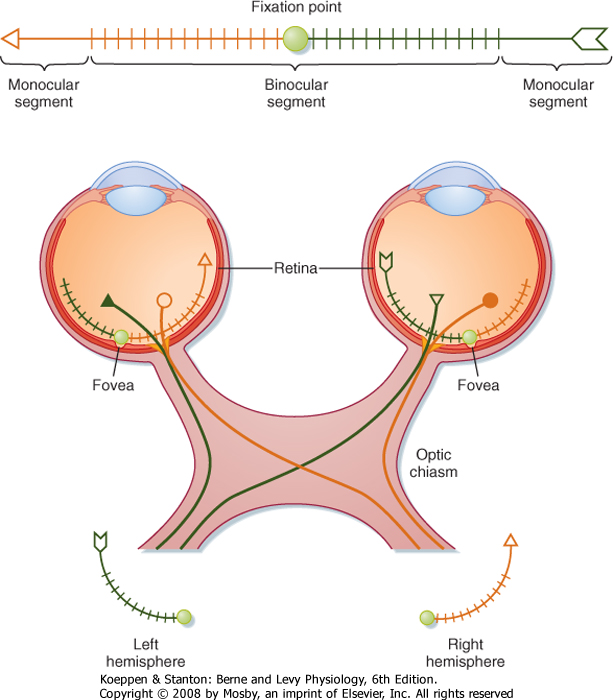
| Retinal ganglion cells transmit information to the brain by way of the optic nerve, optic chiasm, and optic tract. Figure 8-9 shows the relationships between a visual target (arrow), the retinal images of the target in the two eyes, and the projections of retinal ganglion cells to the two hemispheres of the brain. The eyes and the optic nerves, chiasm, and tract are viewed from above.
|
| Figure 8-9 Relationships between a visual target, images on the retinas of the two eyes, and projections of the ganglion cells carrying visual information about these images. The image is so large that it extends into the monocular segments of the eyes where the image is seen in only one eye. Note that all the information about the left visual field of both eyes is conveyed to the right side of the brain and all the information about the right visual field is conveyed to the left side. |
| page 133 | | | page 134 |
| The visual target, an arrow, is in the visual fields of both eyes (Fig. 8-9) and, in this case, is so long that it extends into the monocular segments of each retina (i.e., one end of the target can be seen only by one eye and the other end only by the other eye). The shaded circle at the center of the target shows the fixation point. The image of the target is reversed on the retinas by the lens system. The left half of the visual target is
imaged on the nasal retina of the left eye and the temporal retina of the right eye. Thus, the left visual field is seen by the left nasal retina and the right temporal retina. Similarly, the right half of the visual target is imaged on and seen by the left temporal retina and the right nasal retina. There is also an inversion in the vertical axis, with the upper visual field imaged on the lower retina and vice versa.
|
| The projections of retinal ganglion cells may be uncrossed or crossed, depending on the location of the ganglion cell in the retina (Fig. 8-9). Axons from the temporal portion of each retina pass through the optic nerve, the lateral side of the optic chiasm, and the ipsilateral optic tract and terminate ipsilaterally in the brain. Axons from the nasal portion of each retina pass through the optic nerve, cross to the opposite side in the optic chiasm, and then pass through the contralateral optic tract to end in the contralateral side of the brain. This arrangement results in the representation of objects in the left field of vision in the right side of the brain and those in the right field of vision in the left side of the brain (Fig. 8-10).
|
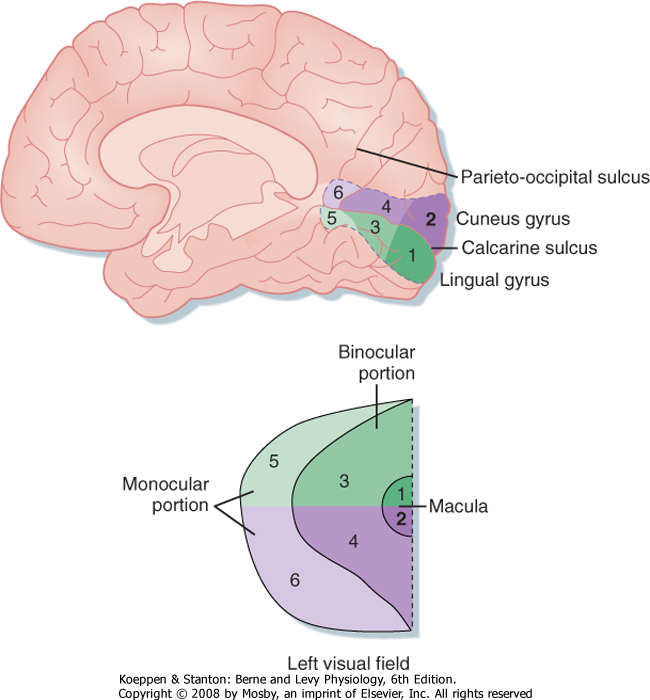
|
| Figure 8-10 The left visual field is relayed, via the LGN and visual radiation to the primary visual cortex of the right hemisphere, as a point-to-point retinotopic map. The representation of each part of visual space is proportional to the number of afferent axons with receptive fields in that part of space. As a result, the area of macular representation (near the occipital pole) is larger than that for the rest of the binocular and monocular fields. Note that the lower half of the field is represented in the cuneus gyrus above the calcarine fissure and the upper half of the field in the lingual gyrus below the fissure. (Redrawn from Purves D et al [eds]: Neuroscience, 3rd ed. Sunderland, MA, Sinauer, 2004.) |
| Retinal ganglion cell axons can synapse in several nuclei of the brain, but the main target for vision is the LGN of the thalamus. The LGN in turn projects to the primary visual cortex or striate cortex by way of the visual radiation. The visual radiation fans out, and the fibers carrying information derived from the lower half of the appropriate hemiretinas (and therefore the contralateral upper visual field) project to the lingual gyrus, which lies on the medial surface of the occipital lobe, just below the calcarine fissure. Axons in the visual radiation that represent the contralateral lower
visual field project to the adjacent cuneus gyrus, which lies just above the calcarine fissure. Together, the portions of these two gyri that line and border the calcarine fissure constitute the primary visual cortex (or area 17) (Fig. 8-10).
|
| Interruption of the visual pathway at any level will cause a defect in the appropriate part of the visual field (Fig. 8-9). For example, a tiny lesion in the retina would result in a blind spot (scotoma) in that eye, whereas a similar lesion in the striate cortex would produce corresponding scotomas in both eyes. Interruption of the optic nerve on one side produces blindness in that eye. Damage to the optic nerve fibers as they cross in the optic chiasm results in loss of vision in both temporal fields of vision; this condition is known as bitemporal hemianopsia and occurs because the crossing fibers originate from ganglion cells in the nasal halves of each retina. A lesion of the entire optic tract, LGN, visual radiation, or visual cortex on one side causes homonymous hemianopsia, which is loss of vision in the entire contralateral visual field. Partial lesions result in partial visual field defects. For example, a lesion in the lingual gyrus causes an upper homonymous quadrantanopsia, which in this case is loss of vision in the contralateral, upper visual field. |
|
| page 134 | | | page 135 |
| In addition, the representation of the macula occupies the most posterior and largest part of both gyri, with progressively more peripheral retina projected to
more anterior parts of these gyri. Overall, there is point-to-point mapping of retinal loci across the surface of the striate cortex.
|
| Lateral Geniculate Nucleus
|
| The LGN is a layered structure. The first two layers, which contain large neurons, are called the magnocellular layers. The other four layers are the parvocellular layers. There is a point-to-point projection from the retina to the LGN. The LGN thus has a retinotopic map. Cells that represent a particular retinal location are aligned along projection lines that can be drawn across the layers of the LGN.
|
| The projection from each eye is distributed to three of the layers of the LGN, one of the magnocellular layers (layers 1 and 2 get M cell input) and two parvocellular layers (layers 3 to 6 get P cell input). Color-coded ganglion cells project to groups of cells between the major layers, the intralaminar zones. Thus, the properties of LGN neurons are very similar to those of retinal ganglion cells. For example, LGN neurons can be classified as P or M cells, and they have on-center or off-center receptive fields.
|
| The LGN also receives input from the visual areas of the cerebral cortex, the thalamic reticular nucleus, and several nuclei of the brainstem reticular formation. The activity of LGN projection neurons is inhibited by interneurons both in the LGN and in the thalamic reticular nucleus. These cells use γ-aminobutyric acid (GABA) as their inhibitory neurotransmitter. In addition, the activity of LGN neurons is influenced by corticofugal pathways and by brainstem neurons that use monoamine transmitters. These control systems filter visual information and may be important for selective attention.
|
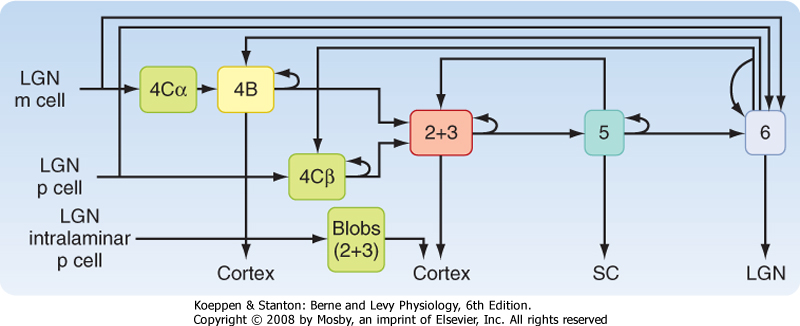
| The geniculostriate pathway ends chiefly in layer 4 of the striate cortex (Fig. 8-11), with the M and P cells segregated into separate sublayers 4Cα and 4Cβ, respectively, while the projection from the intralaminar LGN terminates in so-called blobs in layers 2 and 3. Similarly, axons that represent one eye or the other terminate within layer 4C in alternate patches that define ocular dominance columns. Cortical neurons in such a column respond preferentially to input from one eye. Near the border between two ocular dominance columns, neurons respond about equally to input from the two eyes.
|
| Figure 8-11 Diagram of visual information flow into the visual cortex from the LGN and its projection to the extrastriate cortex. M, magnocellular path; P, parvocellular path. (Redrawn from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.) |
| Like the LGN, the striate cortex contains a retinotopic map (actually, two interlaced retinotopic maps, one for each eye). The macula is represented by a relatively large region in comparison to the remainder of the retina. The macular representation extends forward from the occipital pole for about a third the length of the striate cortex (Fig. 8-10).
|
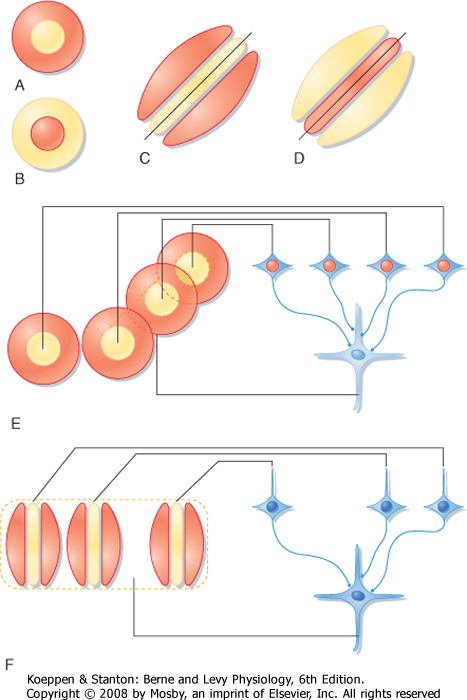
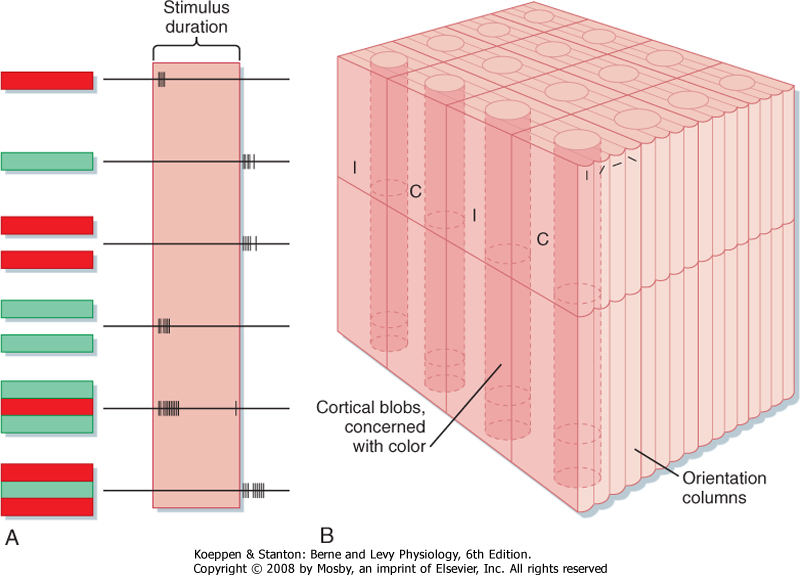
| The receptive fields of neurons in the striate cortex, aside from the monocular cells in layer 4C, are more complex than those of LGN neurons. Neurons in other layers may be binocular and respond to stimulation of both eyes, although the input from one eye often dominates (see Chapter 10). In addition, cortical neurons outside layer 4C often show orientation selectivity (i.e., they respond best when the stimulus, such as a bar or an edge, is oriented and positioned in a particular way) (Fig. 8-12). These "simple cells" appear to be responding as though they received input from cells whose concentric center-surround receptive fields were arranged such that their "on" centers were aligned in a row flanked by antagonistic regions. "Complex" cortical neurons are similar to "simple" cells in that they are orientation specific, but instead of having flanking excitatory and inhibitory zones, they respond best to a particular stimulus orientation anywhere in their receptive field. They may also display direction selectivity; that is, they may respond when the stimulus is moved in one direction but not when it is moved in the opposite direction (Fig. 8-12). The receptive field of a "complex" cell may be thought of as a composite of adjacent "simple" cells with similar orientation selectivity. Because such neurons in a particular zone of the cortex all tend to have the same orientation selectivity, they are considered to form an orientation column (Fig. 8-13).
|
| However, this classification does not take into account the separate P and M cell pathways. Presumably, parallel P and M cell pathways contribute to the complexity of visual cortical organization. Cortical receptive field organization may depend on both serial and parallel processing.
|
| page 135 | | | page 136 |
| Figure 8-12 Simple and complex receptive fields in the visual cortex can be generated from multiple inputs with concentric fields. A and B represent on-center and off-center input. If three on-center cells (A) with adjacent receptive fields converged onto one cortical neuron (E), that neuron, a simple cell, would respond best to a long bar stimulus at a specific location and orientation (C). For three off-center inputs (B), the resulting receptive field is shown in D. The convergence of multiple simple cells onto another cortical neuron (F) would result in a complex cell that responds best to a bar stimulus with a specific orientation that can be placed anywhere within its receptive field. (Redrawn from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.) |
| Stereopsis is defined as binocular depth perception and appears to be dependent on slight differences in the retinal images formed in the two eyes. Such disparities give different perspectives that lead to visual cues about depth. Stereopsis is useful only for relatively nearby objects. Such perception must be a cortical function because it depends on convergent input
from the two eyes. Depth cues are also available when a single eye is used. For example, the brain interprets distance according to the relative size of familiar objects.
|
| As already discussed, color vision may depend on the presence in the retina of three different types of cones, as well as neurons in the visual pathway that show spectral opposition. Retinal ganglion cells, LGN neurons, and some P cells display spectral opponent properties. The spectral opponent neurons in the striate cortex are found in cortical "blobs," and these show double-opponency in which both the center and the surround respond antagonistically to two colors. Such a cell with R+G- in the center and R-G+ in its surround is shown in Figure 8-13, A. The relationships between the ocular dominance and orientation columns and cortical color blobs are shown in Figure 8-13, B.
|
| Extrastriate Visual Cortex
|
| page 136 | | | page 137 |
| Figure 8-13 A, The receptive field and responses of a neuron in the striate cortex that responds to various combinations of red and green bars. The best on response was to a red bar flanked by two green bars. B, Diagram of the columnar arrangement of the visual cortex. Ocular dominance columns are indicated by I (for ipsilateral) and C (for contralateral). Orientation columns are indicated by the smaller columns marked with short bars at various angles. The cortical blobs contain neurons like that of A and have spectral opponent receptive fields. |
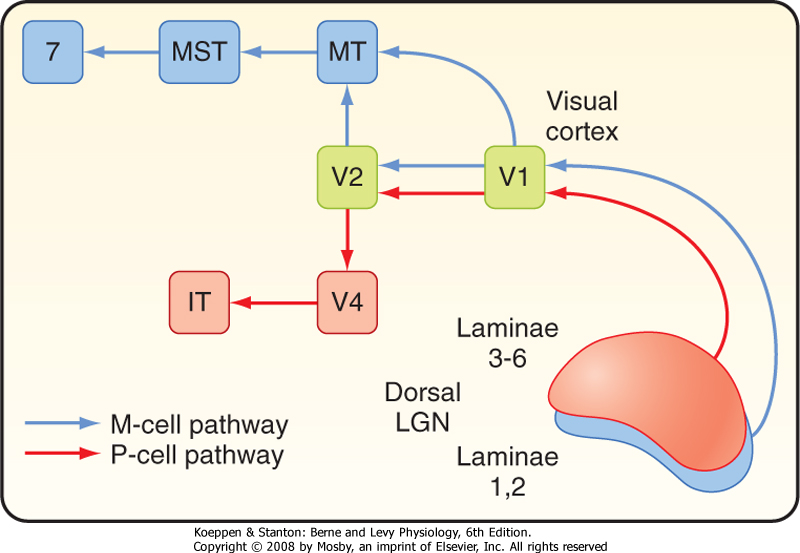
| Figure 8-14 Distribution of P and M cell influences on different areas of the visual cortex. IT, inferotemporal area; MST, medial superior temporal; MT, medial temporal; V1, striate cortex; V2, V4, higher-order visual areas. |
| In animal studies, at least 25 different visual areas have been identified in the cerebral cortex, in addition to the striate cortex (area 17 or V1). The extrastriate areas include several different parallel visual processing pathways. The P pathway originates with P cells and functions in the recognition of form and color. Some of the cortical structures in the P pathway include LGN layers 3 to 6, layer 4Cb of the striate cortex, V4 (Brodmann's area 19), and several areas in the inferotemporal region (Fig. 8-14). Processing of form includes recognition of complex visual patterns, such as faces. Color information is processed separately from form. The M pathway originates with M cells and functions in motion detection and control of eye movement. Cortical structures in the M pathway include layers 4B and 4Ca of the striate cortex and areas MT (medial temporal) and MST (medial superior
temporal) on the lateral aspect of the temporal lobe, as well as area 7a of the parietal lobe (Fig. 8-14). Both P and M pathways contribute to depth perception.
|
| Lesions of the extrastriate visual cortex can produce various deficits. Bilateral lesions of the inferotemporal cortex can result in cortical color blindness (achromatopsia) or in an inability to recognize faces, even of close members of the family (prosopagnosia). A lesion in area MT or MST can interfere with motion detection and eye movements. |
|
| The separation of M and P pathways from the retina through the thalamus and all the cortical regions raises the issue of how all the parts are combined to account for the clear, coherent images of events, objects, and persons that we perceive. It seems unlikely that all the components that represent a percept, such as recognizing a face and then identifying it as belonging to a familiar person, are somehow converged onto a single neuron that will recognize it. The process by which we achieve this "binding" of disparate elements into a percept is unclear, but one working hypothesis
is that it may be accomplished by the temporal synchronization of many anatomically distributed neural events.
|
| The superior colliculus of the midbrain is a layered structure. The three most superficial layers are involved exclusively in visual processing, whereas the deeper layers have multimodal input from the somatosensory and auditory systems, as well as the visual system, particularly from cortical areas involved in eye movement (see Chapter 9).
|
| page 137 | | | page 138 |
| Neurons in the superficial layers of the superior colliculus receive a projection from retinal ganglion cells and are organized into a retinotopic map. Collicular neurons are particularly sensitive to rapid stimulus motion in a particular direction. Most of the cells have binocular input, but they lack orientation selectivity. The ganglion cells include both W and M cells (but not P cells), and they are located chiefly in the contralateral nasal retina. Neurons in the superficial layers of the superior colliculus also receive a projection from the visual cortex, including the striate cortex. The
cortical loop involves neurons activated by M cells. The superficial layers of the superior colliculus, in turn, project to several thalamic nuclei (pulvinar, LGN), and they are indirectly connected to large areas of the visual cortex.
|
| The deep layers of the superior colliculus receive connections from the somatosensory and auditory pathways, in addition to visual input from the superficial layers. Thus, the deep layers of the superior colliculus contain overlaid somatotopic and retinotopic maps, as well as a map of sound in space. For example, an area that receives information about the contralateral visual field will also receive information about sounds that originate from the contralateral auditory space, as well as information about somatic stimuli applied to the contralateral surface of the body. Moreover, the deep layers of the superior colliculus contain a motor map that controls eye and head position. For instance, activation of neurons in the superior colliculus by a visual target causes movement of the eyes to center the visual target on the fovea. In this way the superior colliculus is involved in reflex responses to the appearance of a novel or threatening object in the visual field. Similarly, a sound or a sudden contact with the body will elicit appropriate eye and head movement to enable visualization of the source of the stimulus. The descending pathways include connections to the oculomotor system and to the spinal cord through the tectospinal tract. See Chapter 9 for information on the role of the superior colliculus in eye movements.
|
| Another retinal projection is to the pretectum, which bilaterally activates parasympathetic preganglionic neurons in the Edinger-Westphal nucleus that cause pupillary constriction in the pupillary light reflex. The pretectal areas are also interconnected through the posterior commissure, and thus the reflex causes both ipsilateral (direct) and contralateral (consensual) pupillary constriction when a light is shown in one eye.
|
| The visual pathways also include connections to nuclei that serve functions other than vision. For example, a retinal projection to the suprachiasmatic nucleus of the hypothalamus controls circadian rhythmicity.
|
| THE AUDITORY AND VESTIBULAR SYSTEMS
|
| The peripheral parts of the auditory and vestibular systems share components of the bony and membranous labyrinths, use hair cells as mechanical transducers, and transmit information to the CNS through the vestibulocochlear (CN VIII) nerve. However, the CNS processing and sensory functions of the auditory and vestibular systems are distinct. The function of the auditory system is to transduce sound. This allows us to recognize environmental cues and to communicate with other organisms. The most complex auditory functions are those involved in language. The function of the vestibular system is to provide the CNS with information related to the position and movements of the head in space. Control of eye movement by the vestibular system is discussed in Chapter 9.
|
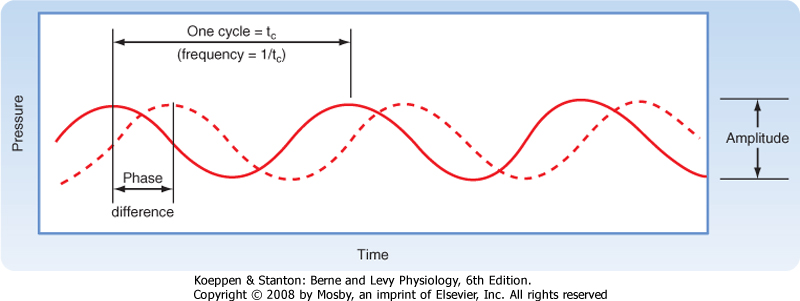
| Figure 8-15 Two pure tones are shown by the solid and dashed lines. Frequency is determined from the wavelength as indicated. Amplitude is the peak-to-peak change in sound pressure. Both tones have the same frequency and amplitude but differ in phase. |
| page 138 | | | page 139 |
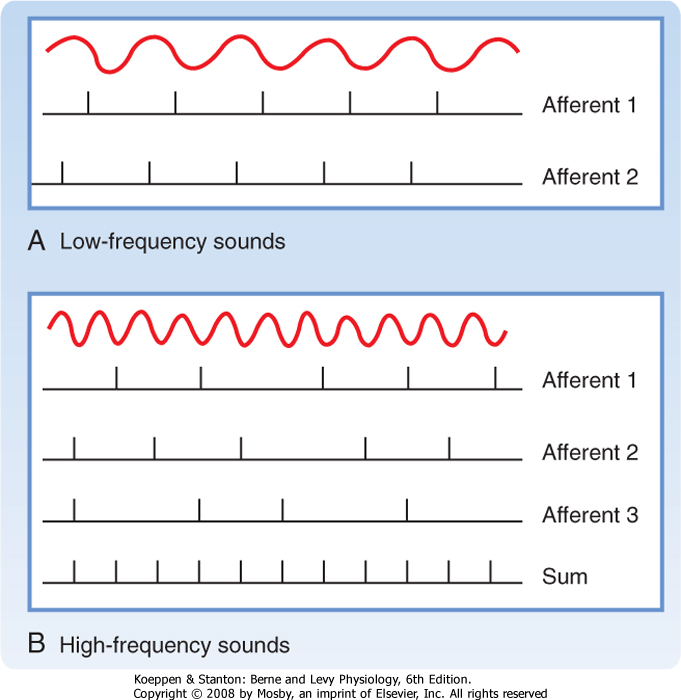
Sound is produced by compression and decompression waves that are transmitted in air or in other elastic media such as water. Sound frequency is measured in cycles per second, or hertz (Hz). Each pure tone results from a sinusoidal wave at a particular frequency, and each pure tone is characterized not only by its frequency but also, instantaneously, by its amplitude and phase (Fig. 8-15). Most naturally occurring sound, however, is actually a mixture of pure tones. Noise is unwanted sound and may have any composition of pure tones. Sound propagates at about 335 m/sec in air. The waves are associated with certain pressure changes, called sound pressure. The unit of sound pressure is N/m2, but sound pressure is more commonly expressed as the sound pressure level (SPL). The unit of SPL is the decibel (dB):
 where P is sound pressure and PR is a reference pressure (0.0002 dyne/cm2, the absolute threshold for human hearing at 1000 Hz). A sound with intensity 10 times greater would be 20 dB; one 100 times greater would be 40 dB.
where P is sound pressure and PR is a reference pressure (0.0002 dyne/cm2, the absolute threshold for human hearing at 1000 Hz). A sound with intensity 10 times greater would be 20 dB; one 100 times greater would be 40 dB.
|
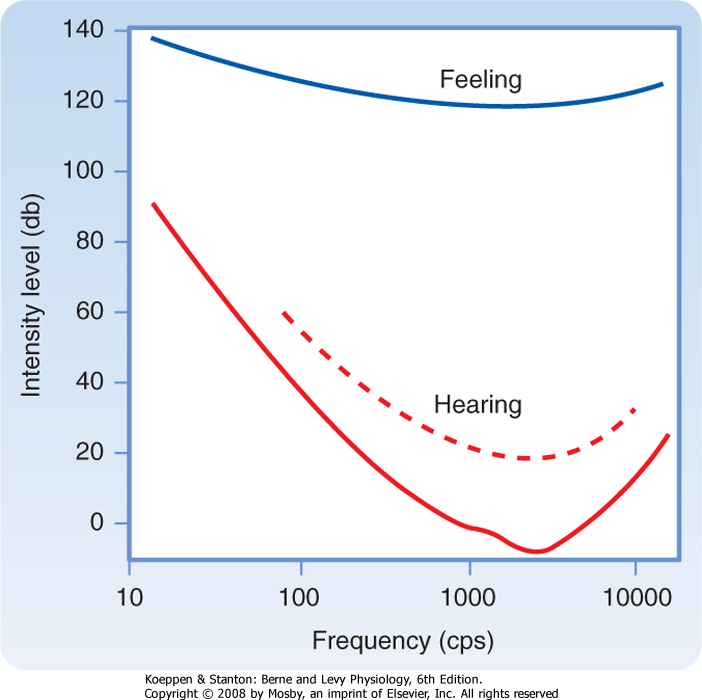
| The normal young human ear is sensitive to pure tones with frequencies that range between about 20 and 20,000 Hz. The threshold for detection of a pure tone varies with its frequency (Fig. 8-16). The lowest thresholds for human hearing are for pure tones around 3000 Hz. The threshold at these frequencies is approximately -3 to -5 dB, as compared with the reference 0 dB at 1000 Hz. According to this scale, speech has an intensity of about 65 dB. The main frequencies used in speech fall in the range of 300 to 3500 Hz. Sounds that exceed 100 dB can damage the peripheral auditory apparatus, and those higher than 120 dB can cause pain and permanent damage. As people age, their thresholds at high frequencies rise, thereby reducing their ability to hear such tones, a condition called presbycusis.
|
| The peripheral auditory apparatus is the ear, which can be subdivided into the external ear, the middle ear, and the inner ear (Fig. 8-17).
|
| Figure 8-16 Sound threshold intensities at different frequencies. The bottom curve indicates the absolute intensity needed to detect a sound. The dashed curve is the threshold for functional hearing. The top curve indicates levels at which sound is felt as painful. |
| The external ear includes the pinna, the external auditory meatus auditory canal. The auditory canal contains glands that secrete cerumen, a waxy protective substance. The pinna helps direct sounds into the auditory canal and plays a role in sound localization. The auditory canal transmits the sound pressure waves to the tympanic membrane. In humans, the auditory canal has a resonant frequency of about 3500 Hz, and this frequency contributes
to the low threshold sensitivity in that range.
|
| The external ear is separated from the middle ear by the tympanic membrane (Fig. 8-17, A). The middle ear contains air. A chain of ossicles connect the tympanic membrane to the oval window, an opening into the inner ear. Adjacent to the oval window is the round window, another membrane-covered opening between the middle and inner ear (Fig. 8-17, A and B).
|
| The ossicles include the malleus, the incus, and the stapes. The stapes has a footplate that inserts into the oval window. Behind the oval window is a fluid-filled component of the cochlea. This component is called the vestibule, and it is continuous with a tubular structure known as the scala vestibuli. Inward movement of the tympanic membrane by a sound pressure wave causes the chain of ossicles to push the footplate of the stapes into the oval window (Fig. 8-17, B). This movement of the stapes footplate in turn displaces the fluid within the scala vestibuli. The pressure wave that ensues within the fluid is transmitted through the basilar membrane of the cochlea to the scala tympani (see later), and it causes the round window to bulge into the middle ear.
|
| The middle ear also serves other functions. Two muscles are found in the middle ear: the tensor tympani attached to the malleus and the stapedius attached to the stapes. When these muscles contract, they damp movements of the ossicles and decrease the sensitivity of the acoustic apparatus. This action can protect the acoustic apparatus against damaging sounds that can be anticipated. However, a sudden explosion can still damage the acoustic apparatus because reflex contraction of the middle ear muscles does not occur quickly enough. The chamber of the middle ear connects to the pharynx through the eustachian tube. Pressure differences between the external and middle ear can be equalized through this passage. If fluid collects in the middle ear, such as during an infection, the eustachian tube may become blocked. The resulting pressure difference between the external and middle ear can produce pain displacement of the tympanic membrane and, in extreme cases, by rupture of the tympanic membrane. Unequalized pressure changes as a result of flying or diving can also cause discomfort. |
|
| page 139 | | | page 140 |
| Figure 8-17 Ear and cochlear structure. A, Location of the right human cochlea in relation to the vestibular apparatus and middle and external ears. B, Relationships between the outer, middle, and inner ear spaces, with the cochlea unrolled for clarity. C, Drawing of a cross section through the cochlea. The organ of Corti (Fig. 8-18A, B) is outlined. |
| page 140 | | | page 141 |
| The tympanic membrane and the chain of ossicles serve as an impedance-matching device. The ear must detect sound waves traveling in air, but the neural transduction mechanism depends on movement in the fluid-filled cochlea, where acoustic impedance is much higher than that of air. Therefore, without a special device for impedance matching, most sound reaching the ear would simply be reflected, as are voices from shore when you are swimming under water. Impedance matching in the ear depends on (1) the ratio of the surface area of the large tympanic membrane to
that of the smaller oval window and (2) the mechanical advantage of the lever system formed by the ossicle chain. This impedance matching is sufficient to increase the efficiency of energy transfer by nearly 30 dB in the range of hearing from 300 to 3500 Hz.
|
| The inner ear includes the bony and membranous labyrinths. The bony labyrinth is a complex, but continuous series of spaces in the temporal bone of the skull, whereas the membranous labyrinth consists of a series of soft tissue spaces and channels lying inside the bony labyrinth. The cochlea and the vestibular apparatus are formed from these structures.
|
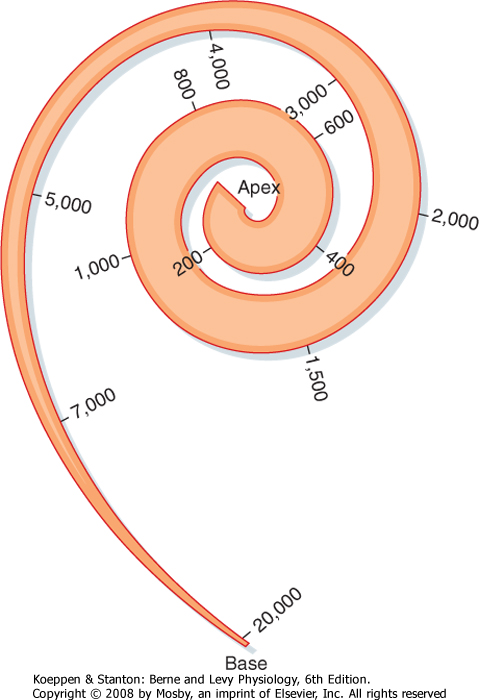
| The cochlea is a spiral-shaped organ (Fig. 8-17, A and B). In humans, the spiral consists of 2¾ turns from a broad base to a narrow apex, although its internal lumen is small at the base and wide at the top. The apex of the cochlea faces laterally (Fig. 8-17, A). The bony core around which the cochlea coils is the modiolus.
|
| The bony labyrinth component of the cochlea is subdivided into several chambers. The vestibule is the space facing the oval window (Fig. 8-17, A). Continuous with the vestibule is the scala vestibuli, a spiral-shaped chamber that extends to the apex of the cochlea, where it meets and merges with the scala tympani at the helicotrema. The scala tympani is another spiral-shaped space that winds back down the cochlea to end at the round window (Fig. 8-17, B). Separating the two, except at the helicotrema, is the scala media enclosed in the membranous labyrinth.
|
| The scala media, or cochlear duct (Fig. 8-17, B and C), is a membrane-bound spiral tube that extends 35 mm along the cochlea, between the scala vestibuli and scala tympani. One wall of the scala media is formed by the basilar membrane, another by Reissner's membrane, and the third by the stria vascularis (Fig. 8-17, C).
|
| The spaces within the cochlea are filled with fluid. The fluid in the bony labyrinth, including the scala vestibuli and scala tympani, is perilymph, which closely resembles cerebrospinal fluid. The fluid in the membranous labyrinth, including the scala media, is endolymph, which is very different from perilymph. Endolymph contains high [K+] (about 145 mM) and low [Na+] (about 2 mM); in this respect it resembles intracellular fluid. Because endolymph has a positive potential (about +80 mV), a large potential gradient (about 140 mV) exists across the membranes of the hair cells found within the cochlea. (These hair cells, which are the sensory receptors for sound, are discussed in more detail later.) Endolymph is secreted by the stria vascularis and is drained through the endolymphatic duct into the dural venous sinuses.
|
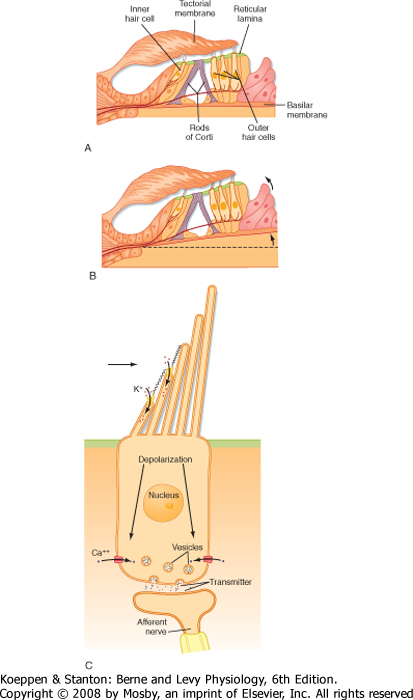
| The neural apparatus responsible for transduction of sound is the organ of Corti (Fig. 8-17, C), which is located within the cochlear duct. It lies on the basilar membrane and consists of several components, including three rows of outer hair cells, a single row of inner hair cells, a gelatinous tectorial membrane, and a number of types of supporting cells. The organ of Corti in humans contains 15,000 outer and 3500 inner hair cells. The rods of Corti help provide a rigid scaffold. Located on the apical surface of the hair cells are stereocilia, which can be described as nonmotile cilia that contact the tectorial membrane.
|
| The organ of Corti is innervated by nerve fibers that belong to the cochlear division of the eighth cranial nerve. The 32,000 auditory afferent fibers in humans originate in sensory ganglion cells in the spiral ganglion, which is located within the modiolus. These nerve fibers penetrate the organ of Corti and terminate at the base of the hair cells (see Fig. 8-17, C and 8-18). About 90% of the fibers end on inner hair cells, and the remainder end on outer hair cells. Thus, in this arrangement about 10 afferent fibers supply each inner hair cell, whereas other afferent fibers diverge to supply about five outer hair cells each. The inner hair cells clearly provide most of the neural information about acoustic signals that the CNS uses for hearing. The function of the outer hair cells is less clear.
|
| In addition to afferent fibers, the organ of Corti is supplied by efferent fibers, most of which terminate on the outer hair cells. These cochlear efferents originate in the superior olivary nucleus of the brainstem and are often called olivocochlear fibers. The length of the outer hair cells varies; this characteristic suggests that changes in outer hair cell length may affect the sensitivity, or "tuning," of the inner hair cells. The cochlear efferent fibers may control outer hair cell length. Such a mechanism could conceivably influence the sensitivity of the cochlea and the way that the brain recognizes sound. Other efferent fibers that end on cochlear afferent fibers may be inhibitory, and they may help improve frequency discrimination.
|
| A common cause of deafness is the destruction of hair cells by loud sounds. Hair cells can be destroyed, for example, by exposure to industrial noise or by listening to loud music. Typically, hair cells in certain parts of the cochlea are selectively damaged, and thus hearing may be lost over a discrete frequency range. Presbycusis, or the loss of high-frequency hearing with age, is probably increased by the loss of hair cells because of long-term noise exposure in urban environments. |
| page 141 | | | page 142 |
| Figure 8-18 A and B, Detail of the area indicated in Figure 8-17, C, showing the organ of Corti and demonstrating how movement of the basilar membrane will cause the stereocilia to bend because of shear forces produced by relative displacement of the hair cells and the tectorial membrane. C, Diagram of a hair cell with tip-link connections between the hair cell cilia to demonstrate how shear forces open mechanoreceptor channels. |
| Sound waves are transduced by the organ of Corti. Sound waves that reach the ear cause the tympanic membrane to oscillate, and these oscillations are transmitted to the scala vestibuli by the ossicles. This creates a pressure difference between the scala vestibuli and the scala tympani (Fig. 8-17, B) that serves to displace the basilar membrane and, with it, the organ of Corti (Fig. 8-18, A and B). Because of the shear forces set up by the relative displacement of the basilar and tectorial membranes, the stereocilia of the hair cells bend. Upward displacement bends the stereocilia toward the tallest cilium (away from the modiolus),
which depolarizes the hair cells; downward deflection bends the stereocilia in the opposite direction, which hyperpolarizes the hair cells.
|
| In view of the wide range of frequencies and amplitudes of sound stimuli, it is no surprise that hair cell transduction must provide for a fast response. The fast response to deflection of the cilia is based on direct opening of ion channels by "tip links" that connect the tip of each stereocilium with the shaft of the next taller one (Fig. 8-18, C). With deflection, the tip links are subjected to a lever action that transiently opens the channels, permits the entry of K+ (because of the high [K+] and high potential in endolymph), and depolarizes the hair cell. Several mechanisms have been proposed to account for the equally important rapid adaptation necessary for a high-frequency response. A "spring" response by the tip links would allow the attachment point of the tip link to be moved along the stereocilium's shaft to reset the mechanical leverage of the tip link. In addition, it has been observed that Ca++ can enter and bind to the open channel, change it to require greater opening force, and thereby reduce the statistical probability of opening.
|
| The potential gradient that induces movement of ions into hair cells includes both the resting potential of the hair cells and the positive potential of the endolymph. As noted previously, the total gradient across the apical membrane of hair cells is about 140 mV. Therefore, a change in membrane conductance in the apical membranes of hair cells results in a rapid current flow that produces the receptor potential in these cells. This current flow can be recorded extracellularly as a cochlear microphonic potential, an oscillatory event that has the same frequency as the acoustic stimulus. The cochlear microphonic potential represents the sum of the receptor potentials of a number of hair cells.
|
| Hair cells, like retinal photoreceptors, release an excitatory neurotransmitter (probably glutamate) when depolarized. The transmitter produces a generator potential in the cochlear afferent nerve fibers with which the hair cell synapses. In summary, sound is transduced when oscillatory movements of the basilar membrane cause transient changes in the transmembrane voltage of the hair cells and, consequently, the generation of action potentials in cochlear afferent nerve fibers. The activity of a large number of cochlear afferent fibers can be recorded extracellularly as a compound action potential.
|
| page 142 | | | page 143 |
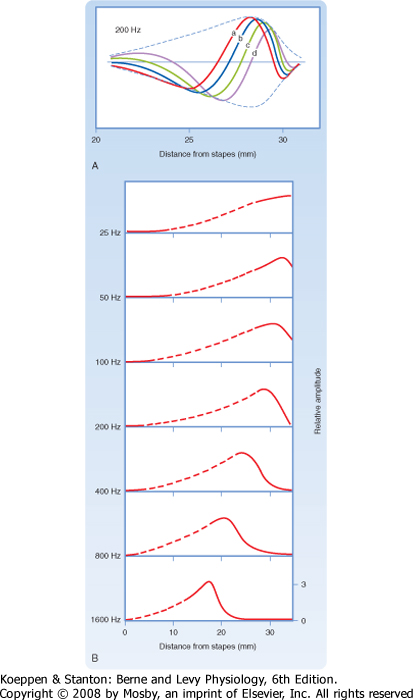
| However, not all the cochlear afferent fibers discharge in response to a particular sound frequency. One factor that influences which afferent fibers discharge is their location along the organ of Corti. The location of an afferent fiber is important because for any given sound frequency, there is a site of maximum displacement as the pressure wave travels along the basilar membrane (Fig. 8-19). The location varies because the width and tension along the basilar membrane vary with distance from the base.
|
| On the basis of these differences in width and tension, investigators originally concluded that different parts of the basilar membrane have different resonance frequencies. For example, the basilar membrane is about 100 μm wide at the base and 500 μm wide at the apex. It also has higher tension at the base. Thus, the base was predicted to vibrate at higher frequencies than the apex, as do the shorter strings of musical instruments. However, experiments have shown that the basilar membrane moves as a whole in traveling waves (Fig. 8-19). Movements of the basilar membrane are maximal nearer the base of the cochlea during high-frequency tones and maximal nearer the apex during low-frequency tones.
|
| In effect, the basilar membrane serves as a frequency analyzer; it distributes the stimulus along the organ of Corti so that different hair cells will respond differentially to particular frequencies of sound. This is the basis of the place theory of hearing. In addition, hair cells located at different places along the organ of Corti are tuned to different frequencies because of differences in their stereocilia and biophysical properties. As a result of these factors, the basilar membrane and organ of Corti have a so-called tonotopic map (Fig. 8-20).
|
| The activity of hair cells in the organ of Corti causes action potentials in the primary afferent fibers of the cochlear nerve. These afferents of the vestibulocochlear CN VIII nerve are bipolar cells with a myelin sheath around the cell bodies, as well as around the axons. Their cell bodies are in the spiral ganglion, their peripheral processes end on hair cells, and their central processes terminate in the cochlear nuclei of the brainstem.
|
|
| Figure 8-19 Different frequencies of sound result in different amplitudes of displacement of the basilar membrane at different sites along the organ of Corti. A, Traveling wave produced in the basilar membrane by a sound of 200 Hz. The curves at a, b, c, and d represent displacement of the basilar membrane at different times, and the dashed line is the envelope formed by the peaks of the wave at different times. Maximum deflection occurs at about 29 mm from the oval window. B, Envelopes of traveling waves produced by several frequencies of sound. Note that the maximum displacement varies with frequency and is closest to the stapes when the frequency is highest. (Redrawn from von Bekesy G: Experiments in Hearing. New York, McGraw-Hill, 1960.) |
| page 143 | | | page 144 |
| Figure 8-20 Layout of the tonotopic map of the cochlea. (Redrawn from Stuhlman O: An Introduction to Biophysics. New York, John Wiley & Sons, 1943.) |
| Characteristic Frequencies
|
| A cochlear afferent fiber discharges maximally when stimulated by a particular sound frequency called the characteristic frequency of that fiber. The characteristic frequency can be determined from a tuning curve for the fiber (Fig. 8-21). A tuning curve plots the threshold for activation of the nerve fiber by different sound frequencies. Typically, tuning curves are sharp near the threshold, but they broaden at high sound pressure levels. Both excitatory and inhibitory areas can be included in a tuning curve (Fig. 8-21, A). The sharpness of some tuning curves may reflect inhibitory processes.
|
| An important, though relatively uncommon, condition that can interrupt the function of cochlear nerve fibers is an acoustic neuroma, a tumor of Schwann cells of the eighth nerve. As the tumor grows, irritation of cochlear nerve fibers may cause a ringing sound in the affected ear (tinnitus). Eventually, conduction in cochlear nerve fibers is blocked, and the ear becomes deaf. The tumor may be operable while still small; therefore, early diagnosis is important. If the tumor is allowed to enlarge substantially, it could interrupt the entire eighth nerve and cause vestibular as well as auditory difficulties. It could also impinge on or distort neighboring cranial nerves (e.g., V, VII, IX, and X), and it could produce cerebellar signs by compressing the cerebellar peduncles. |
|
| The input from each ear is bilaterally represented in the ascending auditory system pathway at the level of the lateral lemniscus and above. Thus, the representation of auditory space is complex, even at the brainstem level. Consequently, unilateral deafness may occur with isolated lesions of the cochlear nuclei or more peripheral structures. Central lesions do not cause unilateral deafness, although they may interfere with overall sensitivity to speech or with sound localization. |
|
| The different features of an acoustic stimulus are encoded in the discharges of cochlear nerve fibers. Duration is signaled by the duration of activity; intensity is signaled both by the amount of neural activity and by the number of fibers that discharge. For low-frequency sounds, the frequency is signaled by the tendency of an afferent fiber to discharge in phase with the stimulus (phase locking, Fig. 8-22, A). Phase locking can also occur for sounds with periods shorter than the absolute refractory period of the afferent fiber. If the tone is much more than 1 kHz, a single fiber cannot discharge with every cycle. The CNS can detect higher-frequency information, however, from the activity of a population of afferent fibers, each of which discharges in phase with the stimulus and which, as a group, signal the frequency of the stimulus (Fig. 8-22, B). This observation is the basis of the frequency theory of hearing.
For still higher frequencies (>5000 Hz), the place theory must dominate, with the CNS interpreting sounds that activate afferent fibers supplying hair cells near the base of the cochlea as being of high frequency. Thus, both the place and the frequency theories are required to explain the frequency coding of sound (duplex theory) across the entire range from 20 to 20,000 Hz.
|
| Cochlear afferent fibers synapse on neurons of the dorsal and ventral cochlear nuclei. These neurons give rise to axons that contribute to the central auditory pathways. Some of the axons from the cochlear nuclei cross to the contralateral side and ascend in the lateral lemniscus, the main ascending auditory tract. Others connect with various ipsilateral or contralateral nuclei, such as the superior olivary nuclei, which project through the ipsilateral and contralateral lateral lemnisci. Each lateral lemniscus ends in an inferior colliculus. Neurons of the inferior colliculus project to the medial geniculate nucleus of the thalamus, which gives rise to the auditory radiation. The auditory radiation ends in the auditory cortex (areas 41 and 42), located in the transverse temporal gyri in the temporal lobe.
|
| page 144 | | | page 145 |
| Figure 8-21 Tuning curves of neurons in the auditory system. Tuning curves can be considered as receptive field plots. A, Tuning curve with excitatory (E) and inhibitory (I) regions. B, Tuning curves for cochlear nerve fibers (upper left), neurons in the inferior colliculus (upper right), trapezoid body (lower left), and medial geniculate nucleus (lower right). (A, Redrawn from Arthur RM et al: J Physiol [Lond] 212:593, 1971; B, redrawn from Katsui Y. In Rosenblith WA [ed]: Sensory Communication. Cambridge, MA, MIT Press, 1961.) |
| Functional Organization of the Central Auditory System
|
| Receptive Fields and Tonotopic Maps
|
| The responses of neurons in several structures that belong to the auditory system can be described by tuning curves (Fig. 8-21, B). By plotting the distribution of the characteristic frequencies of neurons within a nucleus or in the auditory cortex, a tonotopic map may be revealed in which neurons are ordered by their "best" frequencies. Tonotopic maps have been found in the cochlear nuclei, superior olivary complex, inferior colliculus, medial geniculate nucleus, and auditory cortex. A given auditory structure may, in fact, contain several tonotopic maps.
|
| page 145 | | | page 146 |
| Figure 8-22 A, At low frequencies, individual auditory afferents can respond at each cycle to the signal frequency. B, At higher frequencies, each afferent generates an action potential only at certain cycles, limited by its maximum firing frequency. However, the overall population of afferents can still signal stimulus frequency by their aggregate firing frequency. |
| Most auditory neurons at levels above the cochlear nuclei respond to stimulation of either ear (i.e., they have binaural receptive fields). Binaural receptive fields contribute to sound localization. A human can distinguish sounds originating from sources separated by as little as 1 degree. The
auditory system uses several clues to judge the origin of sounds, including differences in the time (or phase) of arrival of the sound at the two ears and differences in sound intensity on the two sides of the head.
|
| These factors provide information about the location of a sound by influencing the activity of neurons in the superior olivary complex. For example, neurons in the medial superior olivary nucleus have medial and lateral dendrites. The synapses on the medial dendrites are largely excitatory, and they originate from the contralateral ventral cochlear nucleus. Those on the lateral dendrites are mostly inhibitory and come from the ipsilateral ventral cochlear nucleus. Differences in the phase of the sound reaching the two ears affect the strength and timing of the excitation and inhibition reaching a particular medial olivary neuron. The activity of that neuron can then provide information about sound localization. The lateral superior olivary nucleus uses differences in the sound intensity that reaches the two ears to provide information about the source of the sound.
|
| Several features of the primary auditory cortex resemble those of other primary sensory areas. Not only are sensory maps, in this case tonotopic maps, present in the auditory cortex, but this cortical region also performs feature extraction. Neurons in the primary auditory cortex form isofrequency columns (in which the neurons in the column have the same characteristic frequency), and they also form alternating columns, known as summation and suppression columns. Neurons in summation columns are more responsive to binaural than to monaural input. Neurons in suppression columns are less responsive to binaural than to monaural stimulation, and accordingly, the response to one ear is dominant. Some neurons are selective for the direction of frequency change.
|
| Bilateral lesions of the auditory cortex have some effect on the ability to distinguish the frequency or intensity of different sounds, and the ability to localize sound and to understand speech is reduced. Unilateral lesions, however, have little effect, especially if the nondominant (for language) hemisphere is involved. Evidently, frequency discrimination depends on activity at lower levels of the auditory pathway, possibly the inferior colliculus.
|
| As already discussed, unilateral deafness can be caused by damage to the peripheral auditory apparatus or to the cochlear nuclei, but not by other CNS lesions. A discrete loss of hearing for particular frequencies can result from damage to a part of the organ of Corti (e.g., by exposure to intense sound, such as particularly loud music or industrial noise). The degree of deafness can be quantified for different frequencies by audiometry. In audiometry, each ear is presented with tones of different frequencies and intensities. An audiogram is plotted to show the thresholds of each ear for representative frequencies of sound. Comparison with the audiogram of normal individuals shows the auditory deficit (in decibels). The pattern of deficit aids in diagnosis of the cause of the hearing loss.
|
| Two simple tests are often used clinically to distinguish the most important types of deafness: conduction loss and sensorineural loss. Conduction hearing loss occurs in disorders of the external ear (e.g., ear canal blocked by cerumen) or middle ear (e.g., rupture of the eardrum). Sensorineural hearing loss reflects disorders of the inner ear, the cochlear nerve, or central connections.
|
| The Weber test is used to evaluate the magnitude of conduction hearing loss. In this test the base of a vibrating tuning fork is placed against the middle of the forehead and the subject is asked to localize the sound. Normally, the sound is not localized to a particular ear. However, if the person has conductive hearing loss (e.g., because of a punctured tympanic membrane, fluid in the middle ear, otosclerosis, or loss of continuity of the ossicular chain), the sound is localized to the deaf ear because it is conducted to the cochlea through bone. The sound is also conducted to the cochlea of the undamaged ear, but bone-conducted sound does not activate the organ of Corti as well as sound conducted normally through the tympanic membrane and ossicle chain. One reason why the sound in the Weber test is not localized to the normal ear may be that hearing in the normal ear is inhibited by the ambient sound level (auditory masking). Conversely, in subjects with sensorineural hearing loss (e.g., because of damage to the organ of Corti, the cochlear nerve, or the cochlear nuclei), the sound is localized to the normal side.
|
| page 146 | | | page 147 |
| In the Rinne test, a vibrating tuning fork is placed against the mastoid process, and the subject is asked
to indicate when the sound dies out. The tuning fork is then held near the external auditory meatus. In normal subjects the sound is again heard because the sound is more effectively transmitted to the cochlea in air (i.e., air conduction > bone conduction). If the conduction mechanism is damaged, the sound is not heard when the tuning fork is held near the external auditory meatus. Bone conduction in this case is better than air conduction. If the hearing loss is sensorineural, the sound is heard again when the tuning fork is placed by the external auditory meatus because with sensorineural hearing loss, the inner ear and cochlear nerve are less able to transmit impulses regardless of whether the sound vibrations reach the cochlea via air or bone. Thus, because air conduction is more effective than bone conduction, the bone conduction pattern seen with sensorineural hearing loss is the same as in a normal ear.
|
| The vestibular system detects angular and linear accelerations of the head. Signals from the vestibular system trigger head and eye movements to stabilize the visual image on the retina and allow the body to make adjustments in posture that maintain balance. The following description of the vestibular system emphasizes the sensory aspects of vestibular function, and it introduces the central vestibular pathways. The role of the vestibular apparatus in motor control is discussed in Chapter 9.
|
| Structure of the Vestibular Labyrinth
|
| The vestibular apparatus, like the cochlea, consists of a component of the membranous labyrinth located within the bony labyrinth. The vestibular apparatus on each side is composed of three semicircular canals and two otolith organs (Fig. 8-23; see also Fig. 8-17, A). These structures are surrounded by perilymph and contain endolymph. The semicircular canals include the horizontal, superior, and posterior canals. The otolith organs include the utricle and the saccule. A swelling called an ampulla is found on each semicircular canal where it joins the utricle. The semicircular canals all connect with the utricle. The utricle joins the saccule through the ductus reuniens. The endolymphatic duct originates from the ductus reuniens, and it ends in the endolymphatic sac. The saccule connects with the cochlea, through which endolymph (produced by the stria vascularis of the cochlea) can reach the vestibular apparatus.
|
| The three semicircular canals on one side are matched with corresponding coplanar semicircular canals on the other side. The horizontal canals on each side of the head correspond, as do the superior canal on one side and the posterior canal on the other side (Fig. 8-23, B). This arrangement allows the sensory epithelia, in corresponding pairs of canals on the two sides, to cooperate in sensing movement of the head in all planes. An important feature of the horizontal canals is that they are in the horizontal plane with respect to the horizon if the head is first tilted down 30 degrees. The utricle is oriented nearly horizontally; the saccule is oriented vertically.
|
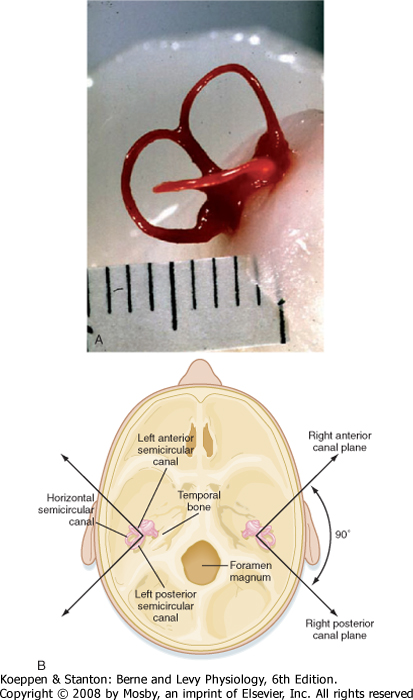
|
| Figure 8-23 A, Lateral view of the right semicircular canals of a Rhesus monkey dissected after filling with plastic. Scale in mm. (Courtesy of Dr. John Simpson, New York University School of Medicine.) B, View of the base of the skull showing the orientation of structures of the inner ear. Coplanar pairs of semicircular canals include the horizontal canals, as well as the superior and contralateral posterior canals. (Redrawn from Haines DE [ed]: Fundamental Neuroscience for Basic and Clinical Applications, 3rd ed. Philadelphia, Churchill Livingstone, 2006.) |
| page 147 | | | page 148 |
| Figure 8-24 Drawing of an ampullary crest inside an ampulla. The stereocilia and the kinocilium of each hair cell extend into the cupula, which extends across the entire cross section of the ampulla. |
| The ampulla of each of the semicircular canals contains a sensory epithelium. The sensory epithelium in a semicircular canal is called a crista ampullaris, or ampullary crest (Fig. 8-24). An ampullary crest consists of a ridge that is covered by epithelium in which
there are vestibular hair cells. These hair cells are innervated by primary afferent fibers of the vestibular nerve, which is a subdivision of the eighth cranial nerve.
|
| Like cochlear hair cells, each vestibular hair cell contains a set of stereocilia on its apical surface. However, unlike cochlear hair cells, vestibular hair cells also contain a large single kinocilium. The cilia on ampullary hair cells are embedded in a gelatinous structure called the cupula. The cupula crosses the ampulla and occludes its lumen completely. Movement of endolymph, produced by angular acceleration of the head in the plane of the canal, deflects the cupula and consequently bends the cilia on the hair cells. The cupula has the same specific gravity as endolymph, and thus it is unaffected by linear acceleratory forces, such as that exerted by gravity.
|
| The sensory epithelia of the otolith organs are called the macula utriculi and the macula sacculi (Fig. 8-25). The hair cells are embedded in the epithelium that overlies each macula. As in the ampullary crests, the stereocilia and kinocilia of the macula project into a gelatinous mass. However, the gelatinous mass in the macula contains numerous otoliths ("ear stones") composed of calcium carbonate crystals. Together, the gelatinous mass and its otoliths are known as an otolithic membrane. The otoliths increase the specific gravity of the otolithic membrane to about twice that of the endolymph. Hence, the otolithic membrane tends to move when subjected to acceleration, whether linear, such as that produced by gravity, or angular, particularly when the center of rotation is outside the head.
|
| Innervation of Sensory Epithelia of the Vestibular Apparatus
|
| The cell bodies of the primary afferent fibers of the vestibular nerve are located in Scarpa's ganglion. The neurons are bipolar, and their cell bodies, as well as axons, are myelinated. The vestibular nerve gives off separate branches to each of the sensory epithelia. The vestibular nerve is accompanied by the cochlear and facial nerves as it enters the internal auditory meatus of the skull.
|
| Like cochlear hair cells, vestibular hair cells are functionally polarized, and the transduction mechanism is presumed to be similar. When the stereocilia are bent toward the longest cilium (in this case, the kinocilium), conductance of the apical membrane increases for cations, and the vestibular hair cell is depolarized (Fig. 8-26). Conversely, when the cilia are bent away from the kinocilium, the hair cell is hyperpolarized. The hair cell releases an excitatory neurotransmitter (either glutamate or aspartate) tonically, so the afferent fiber on which it synapses has a resting discharge. When the hair cell is depolarized, more transmitter is released, and the discharge rate of the afferent fiber increases. Conversely, when the hair cell is hyperpolarized, less transmitter is released, and the firing rate of the afferent fiber slows or stops.
|
| page 148 | | | page 149 |
| Figure 8-25 Structure of the otolith organs. The saccule is shown in A and the utricle in B. (Redrawn from Lindeman HH: Adv Otorhinolaryngol 20:405, 1973.) |
| Angular accelerations of the head produce minute movement of the endolymph in relation to the head (Fig. 8-27). This happens because the inertia of the endolymph causes it to shift in relation to the wall of the membranous labyrinth. This lag distorts the cupula, causes the cilia to bend, and consequently changes the discharge rates of the
vestibular afferent fibers. All the cilia in a given ampullary crest are oriented in the same direction. In the horizontal canal, the cilia are oriented toward the utricle, and in the other ampullae they are oriented away from the utricle.
|
| Irritation of the vestibular labyrinth, as in Meniere's disease, can result in rhythmic conjugate deviations of the eyes, followed by quick return saccades. This condition is known as nystagmus (see Chapter 9). These eye movements are accompanied by a sense of vertigo and often nausea. The brain interprets a difference in the input from the two sides of the vestibular system as head motion. Irritation (or destruction) of one labyrinth produces an asymmetry of input that results in abnormal eye movement and associated psychological effects. |
|
| The way in which angular acceleration of the head affects the discharge of vestibular afferent fibers can
be exemplified by the activity that originates from the horizontal canals. Figure 8-27 shows the horizontal canals and utricle, as seen from above. The hair cells in these canals are polarized toward the utricle. Thus, movement of the endolymph and cilia toward the utricle increases the discharge rate of the afferent fibers, and conversely, movement of the endolymph and cilia away from the utricle reduces the discharge rate.
|
| In Figure 8-27, the head is rotated to the left. As acceleration to the left begins, inertia causes the endolymph in the horizontal canals to increase pressure toward the right. This bends the cilia on hair cells of the ampulla of the left horizontal canal toward the utricle and bends the cilia of the right canal away from the utricle. These actions increase the firing rate in the afferent fibers on the left and decrease the firing rate of the afferent fibers on the right. At a constant velocity of rotation (i.e., no acceleration), there would be no force on either cupula, and therefore the hair cells of both canals would be firing as at rest and at the same rate. However, when the indicated rotation is stopped, the inertia of the endolymph creates a force on both cupulas, but in the opposite direction. This results in an increase in the discharge rate of afferent fibers on the right side and a decrease in the discharge rate on the left. This postrotatory effect is of functional and clinical significance.
|
| page 149 | | | page 150 |
| Figure 8-26 Functional polarization of vestibular hair cells. When the stereocilia are bent toward the kinocilium, the hair cell is depolarized and the afferent fiber is excited. When the stereocilia are bent away from the kinocilium, the hair cell is hyperpolarized and the afferent discharge slows or stops. (Redrawn from Kandel ER, Schwartz JH: Principles of Neural Science. New York, Elsevier, 1981.) |
| Figure 8-27 Effect of head movement to the left on the activity of vestibular afferent fibers supplying hair cells in the horizontal semicircular canals. Small arrows indicate the functional polarity of the hair cells. The large arrow, top, indicates movement of the head; open arrows indicate relative movement of the endolymph. |
| Figure 8-28 Functional polarization of hair cells in the otolith organs. A, The saccule. B, The utricle. The striola in each case is indicated by the dotted line. (Redrawn from Spoendlin HH. In Wolfson RJ [ed]: The Vestibular System and Its Diseases. Philadelphia, University of Pennsylvania Press, 1966.) |
| page 150 | | | page 151 |
| The hair cells in the otolith organs, unlike those in the ampullary crests, are not all oriented in the same direction. Instead, they are oriented with respect to a ridge, called the striola, along the otolith organ (see Fig. 8-25). In the utricle the hair cells on either side of the striola are polarized toward the striola, whereas in the saccule they are polarized away from the striola. Because the striola in each otolith organ is curved, there are hair cells with orientations in all directions (Fig. 8-28). When the head is tilted so that gravity produces a different linear acceleration, the otolithic membranes shift and the cilia of the hair cells bend in a new way. This bending of the cilia of the hair cells changes the pattern of input from the otolith organs to the CNS. Similarly, a linear acceleration caused by other forces, such as might occur in a
free fall or the angular acceleration when turning a car around a curve (angular accelerations have linear centripetal and instantaneous tangential components), will also affect output from the otolith organs.
|
| Central Vestibular Pathways
|
| The vestibular afferent fibers project to the brain-stem through the vestibular nerve. As previously mentioned, the cell bodies of these afferent fibers are located in Scarpa's ganglion. The afferent fibers terminate in the vestibular nuclei, which are located in the rostral medulla and caudal pons. Afferent fibers from different parts of the vestibular apparatus end in different vestibular nuclei and also give off collaterals to the cerebellum.
|
| The vestibular nuclei give rise to various projections, including projections through the medial longitudinal fasciculus to the oculomotor nuclei. Therefore, it is not surprising that the vestibular nuclei exert powerful control over eye movements (the vestibuloocular reflex). Other projections give rise to the lateral and medial vestibulospinal tracts, which provide, respectively, for the activation of trunk and neck muscles and thereby contribute to equilibrium and to head movements (vestibulocolic reflex). There are vestibular projections to the cerebellum, the reticular formation, and the contralateral vestibular complex, as well as to the thalamus. The latter mediate conscious sensation of vestibular activity. Vestibular efferent fibers also originate from the vestibular nuclei. Vestibular reflexes and clinical tests of vestibular function are described in Chapter 9.
|
| The senses of gustation (taste) and olfaction (smell) depend on chemical stimuli that are present either in food and drink or in the air. In the evolution of humans, these chemical senses did not have the survival value of some of the other senses, but they contribute considerably to quality of life and are important stimulants of digestion. In other animals, the chemical senses have greater survival value, and their activation evokes a number of social behaviors, including mating, territoriality, and feeding.
|
| The stimuli that we know as tastes or flavors are actually mixtures of five elementary taste qualities: salty, sweet, sour, bitter, and umami. Taste stimuli that are particularly effective in eliciting these sensations are, respectively, sodium chloride, sucrose, hydrochloric acid, quinine, and monosodium glutamate. Umami has been described as a proteinaceous, meaty flavor.
|
| The sensation of taste depends on the activation of chemoreceptors located in taste buds. A taste bud consists of a group of 50 to 150 receptor cells, as well as supporting cells and basal cells (Fig. 8-29, A). The chemoreceptor cells synapse at their bases with primary afferent nerve fibers. Two types of chemoreceptor cells can be distinguished by differences in their synaptic vesicle content: one type has dense core vesicles, whereas the other has clear round vesicles. The apices of the cells have microvilli that extend toward a taste pore. Chemoreceptor cells live only about 10 days. They are continuously replaced by new chemoreceptor cells that differentiate from basal cells located near the base of the taste bud.
|
| Chemoreceptor molecules, each specialized for one type of taste stimulus, sit on the microvilli of chemoreceptor cells and detect stimulatory molecules that diffuse into the taste pore from the overlying fluid layer. Part of this fluid originates from glands adjacent to the taste buds. Some stimuli can pass directly into the cell to depolarize it (Na+ and H+ for salty and sour) or open cation channels to generate a receptor potential (also salty and sour), whereas others (sucrose, quinine, and glutamate for sweet, bitter, and umami) activate a second messenger that can either open cation channels or directly activate intracellular Ca++ stores (Fig. 8-29, B). In each case, depolarization of the receptor evokes a generator potential that results in the release of an excitatory neurotransmitter and, consequently, action potentials in the primary afferent nerve fiber that are transmitted to the CNS.
|
| Coding of taste, however, is not based entirely on the selectivity of the chemoreceptors for the different primary qualities because each cell responds to a range of stimuli, though most intensely to one. Since most natural flavors have chemicals that effect responses from a number of chemoreceptors, recognition of taste quality appears to depend on the patterned input from a population of chemoreceptors, each responding differentially to the components of the stimulus. The intensity of the stimulus is reflected in the total amount of activity evoked.
|
| Distribution and Innervation of Taste Buds
|
| Taste buds are located on different types of taste papillae found on the tongue, palate, pharynx, and larynx. Types of taste papillae include fungiform and foliate papillae on the anterior and lateral aspects of the tongue and circumvallate papillae on the base of the tongue (Fig. 8-29, C). The latter may contain several hundred taste buds. The tongue in humans may have several thousand taste buds. The sensitivity of different regions of the tongue for different taste qualities varies only slightly because taste buds responding to each type of flavor are widely distributed. The taste buds are innervated by three cranial nerves. The chorda tympani branch of the facial nerve (CN VII) supplies taste buds on the anterior two thirds of the tongue, and the glossopharyngeal nerve (CN IX) supplies taste buds on the posterior third of the tongue (Fig. 8-29, C). The vagus nerve (CN X) supplies a few taste buds in the larynx and upper esophagus.
|
| page 151 | | | page 152 |
| Figure 8-29 Taste bud. A, A taste bud is shown with the taste pore at the top and its innervation below. B, basal cells; R, ciliated taste receptor cells; S, supporting cells. B, Taste receptor cell showing second messenger, ligand-gated, and direct depolarization resulting in depolarization of the cell. C, Distribution of the taste buds on the tongue and their innervation. (Redrawn from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.) |
| page 152 | | | page 153 |
| Figure 8-30 Olfactory chemoreceptors and supporting cells. (Redrawn from de Lorenzo AJD. In Zotterman Y [ed]: Olfaction and Taste. Elmsford, NY, Pergamon, 1963.) |
| Taste is not evaluated in a routine neurological examination. However, a detailed examination can include the application of test substances to the anterior two thirds and posterior third of the tongue on each side. The tongue must be kept protruded to prevent mixing of the test substances with saliva and subsequent redistribution to other areas of the tongue. Taste can also be tested by the application of a galvanic current to the tongue. Taste sensation can be lost, for example, after damage to a cranial nerve that contains gustatory afferents. |
|
| The cell bodies of taste fibers in cranial nerves VII, IX, and X are located in the geniculate, petrosal, and nodose ganglia, respectively. The central processes of the afferent fibers enter the medulla, join the solitary tract, and synapse in the nucleus of the solitary tract.
In some animals, including several rodent species, the second-order taste neurons of the solitary nucleus project rostrally to the ipsilateral parabrachial nucleus. The parabrachial nucleus then projects to the small-celled (parvocellular) part of the ventroposterior medial (VPMpc) nucleus of the thalamus. In monkeys, the solitary nucleus projects directly to the VPMpc nucleus. The VPMpc nucleus is connected to two different gustatory areas of the cerebral cortex, one in the face area of the S1 cortex and the other in the insula. An unusual feature of the central gustatory pathway is that it is predominantly an uncrossed
pathway (unlike the central somatosensory, visual, and auditory pathways, which are predominantly crossed).
|
| The sense of smell is much better developed in animals (macrosmatic animals) than in humans and other primates (microsmatic animals). The ability of dogs to track other animals on the basis of odor is legendary, as is the use of pheromones by insects to attract mates. However, olfaction contributes to our emotional life, and odors can effectively call up memories. In addition, it helps us avoid consuming spoiled food and lets us detect dangerous situations, for example, the strong odorant added to odorless, colorless natural gas.
|
| The olfactory chemoreceptor cells are located in the olfactory mucosa, a specialized part of the nasopharynx. Olfactory chemoreceptors are bipolar nerve cells (Fig. 8-30). The immobile cilia on the apical surface of these cells contain chemoreceptors that detect odorant chemicals dissolved in the overlying mucus layer. From the basal surface, the cell gives off an unmyelinated axon that joins other olfactory nerve filaments and penetrates the base of the skull through openings in the cribriform plate of the ethmoid bone. These olfactory nerves synapse in the olfactory bulb, a portion of the cerebral hemisphere of the brain located at the base of the cranial cavity, just below the frontal lobe (Fig. 8-31).
|
| page 153 | | | page 154 |
| Figure 8-31 Drawing of a sagittal section through an olfactory bulb showing terminations of the olfactory chemoreceptor cells in the olfactory glomerulus and the intrinsic neurons of the olfactory bulb. The axons of the mitral cells are shown exiting in the olfactory tract to the right. (Modified from House EL, Pansky B: A Functional Approach to Neuroanatomy, 2nd ed. New York, McGraw-Hill, 1967.) |
| Humans have about 10 million olfactory chemoreceptors. Like taste cells, olfactory chemoreceptors have a short life span (about 60 days), and they are also continuously replaced. However, olfactory
receptor cells are true neurons and, as such, are the only neurons that are continuously regenerated throughout life.
|
| Odorant molecules are introduced to the olfactory mucosa by ventilatory air currents or from the oral cavity during feeding. Sniffing increases the influx of odorants. The odorants are temporarily bound in mucus to an olfactory binding protein that is secreted by a gland in the nasal cavity.
|
| Odor has more primary qualities than taste does. As many as 1000 different odor receptors are coded in the human genome, and although we probably have only about 350 functional types, they represent the largest population of G protein-coupled receptors in the genome. The olfactory mucosa also contains somatosensory receptors of the trigeminal nerve. When performing clinical tests of olfaction, it is necessary to avoid activating these somatosensory receptors with thermal or noxious stimuli, such as the ammonia used in "smelling salts."
|
| Olfactory coding resembles taste coding in that most natural odors consist of molecules that excite olfactory chemoreceptors of more than one odorant class. Coding for a particular perceived odor depends on the responses of many olfactory chemoreceptors, and the strength of the odorant is represented by the overall amount of afferent neural activity.
|
| The initial relay of the olfactory pathway is located in the olfactory bulb, which is a specialized portion of the cerebral cortex. It contains mitral cells and interneurons (granule cells; periglomerular cells) (Fig. 8-31). The dendrites of mitral cells are long, and they branch to form the postsynaptic components of the olfactory glomeruli. The olfactory afferent fibers that reach the olfactory bulb from the olfactory mucosa ramify as they approach the olfactory glomeruli and synapse on the dendrites of mitral cells. Each glomerulus is the target of thousands of olfactory afferents, each of which share the same type of olfactory receptor. This is all the more remarkable because olfactory receptor cells are being regenerated continuously and new axons must therefore navigate their way to a correct glomerulus. The granule and periglomerular cells are inhibitory interneurons. They form dendrodendritic reciprocal synapses with the dendrites of mitral cells. Activity in a mitral cell depolarizes the inhibitory cells that synapse with it, and they in turn release GABA, an inhibitory neurotransmitter, back into the original and adjacent glomeruli. Because each glomerulus is specialized by being the target of specific olfactory afferents for a unique combination of odor qualities, this appears to be a way of enhancing stimulus contrast, much the way horizontal cells do in the retina. In addition, it provides a mechanism for adaptation to continuous stimulation.
|
| The axons of mitral cells leave the olfactory bulb and enter the olfactory tracts. From here, the olfactory connections become highly complex. Within the olfactory tracts is a nucleus, called the anterior olfactory nucleus, that receives input from the olfactory bulb and projects to the contralateral olfactory bulb through the anterior commissure. As each olfactory tract approaches the base of the brain, it splits into the lateral and medial olfactory striae. Axons of the lateral olfactory stria synapse in the primary olfactory cortex, which includes the prepiriform cortex (and, in many animals, the piriform lobe). The medial olfactory stria includes projections to the amygdala, as well as to the basal forebrain. These structures are portions of, or directly connected to, the limbic system (See Chapters 10 and 11).
|
| page 154 | | | page 155 |
| Olfaction is not generally examined in a routine neurological examination. However, smell can be tested by having the patient inhale and identify an odorant. One nostril should be examined at a time while the other nostril is occluded. Strong odorants, such as ammonia, should be avoided because they also activate trigeminal nerve fibers. Smell sensation can be lost (anosmia) after a basal skull fracture or after damage to one or both olfactory bulbs or tracts by a tumor (such as an olfactory groove meningioma). An aura of a disagreeable odor, often the smell of burning rubber, occurs during uncinate fits, which are epileptic seizures that originate in the medial temporal lobe. |
|
| Note that the olfactory pathway is the only sensory system that does not have an obligatory synaptic relay in the thalamus before reaching the cortex. However, olfactory information does reach the mediodorsal nucleus of the thalamus, and it is then transmitted to the prefrontal and orbitofrontal cortex. The functional roles of olfaction, in addition to the conscious perception of odor, include providing much of the subtleties of taste by enhancing the narrow range of gustatory receptors with the wide repertoire of olfactory receptors. In addition, via its intimate connections with limbic and, by extension, hypothalamic structures, it provides input to subconscious mechanisms related to emotions, memory, and sexual behavior.
|
| page 155 | | | page 156 |
- Light enters the eye through the cornea and lens and is focused on the retina, which lines the back of the eye. The cornea is the most powerful refractive surface, but the lens has a variable power that allows images of near objects to be focused on the retina. The iris regulates depth of field and the amount of illumination that enters the eye.
- The retina has 10 layers. The outermost photoreceptor layer transduces light. Photoreceptors synapse on retinal bipolar cells, which in turn synapse on other interneurons and on ganglion cells. The ganglion cells project to the brain through the optic nerve. The optic disc, where the optic nerve leaves the retina, contains no photoreceptors and is therefore a blind spot. The portion of the retina with the highest degree of spatial resolution is the fovea and the surrounding macula.
- Rod photoreceptors have high sensitivity, no color discrimination, and function best under low light levels. Cone photoreceptors have lower sensitivity but higher spatial resolution. Color vision relies on the three types of cones having different spectral sensitivities.
- Bipolar cells and many ganglion cells have receptive fields with an on-center/off-surround or off-center/on-surround organization. The horizontal cells mediate the center-surround antagonism. Photoreceptor, bipolar, and horizontal cells modulate their membrane potential in response to stimulation, but ganglion cells generate action potentials.
- The axons of ganglion cells in the temporal retina project to the brain ipsilaterally; those in the nasal retina cross in the optic chiasm. The result is that, because the lens inverts the image that falls on the retina, each side of the visual field, from both eyes, is projected to the contralateral side of the brain. In the lateral geniculate nucleus (LGN) of the thalamus, the input from each eye terminates in separate layers, and the M ganglion cells (sensitive to motion) and P ganglion cells (sensitive to detail and color) project to separate layers as well.
- The LGN projects to primary visual (striate) cortex via the visual radiation and terminates largely in layer 4, where there is an orderly retinotopic map. Within the map, information from one or the other eye terminates at adjacent points to create ocular dominance columns that extend vertically in the cortex. Striate cortical neurons outside of layer 4 respond best to bars or edges oriented in a particular way. Cells that prefer a particular stimulus orientation are grouped in orientation columns.
- The many cortical extrastriate visual areas have different functions. Some in the inferotemporal cortex are influenced chiefly by P cells, and they function in form detection, color vision, and face discrimination. M cells influence regions of the middle temporal and parietal cortex, which function in motion detection and the control of eye movements.
- A pure tone is characterized in terms of its amplitude, frequency, and phase. Natural sounds are combinations of pure tones. The unit of sound pressure is the decibel.
- The pinna and auditory canal convey airborne sound waves to the tympanic membrane. The three small bones (ossicles) of the middle ear transmit the vibrations of the tympanic membrane to the oval window of the fluid-filled inner ear. Hearing is most sensitive at about 3000 Hz because of the dimensions of the auditory canal and the mechanics of the ossicles.
- The cochlea of the inner ear has three main compartments: the scala vestibuli, the scala tympani, and the scala media (cochlear duct). The cochlear duct is bounded on one side by the basilar membrane, on which lies the organ of Corti, the sound transduction mechanism.
- When the basilar membrane oscillates in response to pressure waves introduced into the scala vestibuli at the oval window, the stereocilia of the hair cells of the organ of Corti are subjected to shear
forces, which open mechanoreceptor channels. This results in a membrane conductance change that creates a generator potential in cochlear nerve fibers.
- High-frequency sounds best activate the hair cells near the base of the cochlea, and low-frequency sounds activate cells near the apex. Such a tonotopic organization is also found in central auditory structures, including the cochlear nuclei, superior olivary complex, inferior colliculus, medial geniculate nucleus, and primary auditory cortex.
- Auditory processing at many sites in the central auditory pathway contributes to sound localization, frequency and intensity analysis, and speech recognition.
- The vestibular apparatus is part of the inner ear. It includes three semicircular canals (horizontal, superior, and posterior) and two otolith organs (utricle and saccule) on each side. These transduce, respectively, angular and linear accelerations of the head. The three semicircular canals are mutually orthogonal, so they can resolve head acceleration in any plane of motion.
- In each semicircular canal, there are sensory hair cells whose cilia extend into a cupula, which blocks the cross-section of the endolymph-filled canal. Angular head acceleration displaces the endolymph and the cupula, bending the cilia. If the stereocilia bend toward the kinocilium, the hair cell is depolarized, causing an increase in the firing rate in the afferent fiber.
- In the otolith organs, the cilia project into an otolithic membrane. Acceleration of the head, such as with linear movement or change in position with respect to gravity, displaces the otolithic membrane (because of the mass of the otoliths) and changes the firing patterns of the hairs cells, depending on their orientation.
- Central vestibular pathways include afferent connections to the vestibular nuclei and the cerebellum. Activation of the vestibular afferents is detected by the brain as head acceleration or position change and is relayed via the vestibular nuclei to pathways that mediate compensatory eye movements, neck movements, and adjustments to posture.
- Taste buds contain chemoreceptor cells arranged around a taste pore. Taste buds are located on several kinds of papillae on the tongue and in the pharynx and larynx. Five types of taste-receptor cells detect the five elementary qualities of taste: salty, sweet, sour, bitter, and umami. Complex flavors are signaled by patterned population codes using multiple classes of receptor and by central correlation with accompanying olfactory input.
- Afferent taste fibers synapse in the nucleus of the solitary tract. The thalamic relay is to a part of the VPM nucleus, and the taste-receiving areas are located in the SI cortex and the insula.
- Odors are detected by olfactory chemoreceptor cells, which are continuously regenerated in the olfactory mucosa. These cells are true neurons, which are endowed with a wide array of G-protein-coupled receptors that permit the detection of hundreds of odor molecules.
- Individual olfactory axons project to olfactory glomeruli, specific for each stimulus type, in the olfactory bulb. They synapse on the dendrites of mitral cells, which have reciprocal synapse with inhibitory interneurons. This synaptic organization in the glomerulus underlies stimulus adaptation and contrast enhancement.
|
|
|