Acta Botanica Hungarica 31 (1–4), pp. 269–282 (1985) – Akadémiai Kiadó, Budapest
Tankönyvi hivatkozás:
Endrédi Lajos: Biológiai vizsgálatok, 429. oldal – Nemzeti Tankönyvkiadó, Budapest, 2000
EFFECT OF THE SPECTRAL COMPOSITION OF LIGHT ON THE PHYTOMASS PRODUCTION OF LOESS-SWARD,
IN RELATION TO THE CHLOROPHYLL CONTENT
L. Endrédi
TECHERS' TRAINING COLLEGE. DEPARTMENT FOR NATURAL SCIENCES, SZEKSZÁRD, HUNGARY
The effect of the spectral composition of light on the net aboveground phytomass production and the chlorophyll content of dominant species in stands of natural composition of four subassociations of the Salvio-Festucetum rupicolae pannonicum Zólyomi (1958) loess-sward plant association was studied in photostat under controlled conditions. Three kinds of light treatment were used: the plant stands were illuminated by fluorescent lamps with blue, red and compound "white" light respectively, of identical energy content (60 W · m–2) for 12 hours a day over 120 days.
The aboveground drymatter production and energy consumption of the plants significantly varied with the spectral composition of light. In the whole period of examination the highest net production was obtained in red light (6.7032 g ∙ dm–2) and the lowest in blue light (2.3466 g · dm–2) on the average; with plants grown in compound "white" light the dry-matter production (mean value in 120 days: 3.6132 g ∙ dm–2) was closer to that attained in blue light. The energy utilization relatíve to the total radiation durrog the whole period of examination was 3.72 per cent with the red 1.30 per cent with the blue and 2.01 per cent with the compound "white" light.
Changes in the spectral composition of light did not cause essential differences in the average chlorophyll content of the species. For the dominant species a very close positive linear regression correlation between the amount of aboveground phytobiomass and its chlorophyll content on dry weight basis was pointed out in each light treatment. Our results prove the dependence of the production and energy utilization of the vegetation on the number of photons that the chlorophyll molecules are able to absorb.
Introduction
The effect of the spectral composition of light on the intensity of carbon dioxide assimilation, the primary organic matter production and on the products of photosynthesis, respectively, has so far been studied only in certain plant species. Results of experiments with algae were published e.g. by Hauschild, Nelson and Krotkov (1961, 1964), Kowallik (1962), Pirson and Kowallik (1964). Most of the authors studied the photosynthetic production of angiospermous plants (Voskresenskaya and Grishina 1959, Kwack and Dunn 1961, Tregunna, Krotkov and Nelson 1962, Horváth 1965, .Steyenson and Dunn 1965, Koltay and Horváth 1967, Szász 1967, Kadman-Zahayi and Alvarez-Vega 1968, Warrington and Mitchell 1976, Horváth, Mihalik and Takács 1978, Horváth and Mihalik 1980, and others). Many summarizing evaluations have also been published in the subject, e.g. by Veen and meijer (1959), Kleschnin (1960), Nuernbergk (1961), Voskresenskaya (1965, 1972) and Horváth, Mihalik and Takács (1980). We attempted to find out the effect of the spectral composition of light on the net primary production in stands of natural composition of a plant association, under controlled conditions.
We studied the effects of various spectral compositions on the chlorophyll content in the dominant species, and searched for a correlation between the chlorophyll content and the amount of aboveground phytobiomass. The relationship between net production and chlorophyll content has been studied by a number of authors in terrestrial plant associations too [see e.g. the comprehensive work of Fekete (1972). On the correlation in loess-sward vegetation we too have published data (Endrédi and Horváth1977).
The experiment under discussion was carried out with stands of four subassociations of a loess-sward plant association (Salvio-Festucetum rupicolae pannonicum Zólyomi 1958) previously studied first in the field then under controlled conditions, using compound "white" light of 120 W · m–2 energy content (Endrédi and Horváth 1976, 1977, Endrédi 1984). Of the results account was given in two lectures (Endrédi and Horváth 1978, Endrédi1981).
Present paper shows the trend of net aboveground drymatter production and energy consumption in response to different light treatments, with the chlorophyll contents of the dominant species taken into consideration. Data on the photosynthetic pigment contents of the species will be given in detail in another paper.
Material and method
The loess-sward subassociation studied were (Soó 1964, 1973, 1980):
1. Salvio-Festucetum rupicolae panuonicum Zólyomi 1958 festucetosum rupicolae;
2. Salvio-Festucetum rupicolae pannonicum Zólyomi 1958 stipetosum capillatae Soó 1959;
3. Salvio-Festucetum rupicolae pannonicum Zólyomi 1958 andropogonetosum (Boros 1953) Soó 1959;
4. Salvio-Festucetum rupicolae pannonicum Zólyomi 1958 poëtosum angustifoliae Zólyomi 1959.
The stands come from the loess-sward vegetation of the western slope of Nagyhegy at Dunaföldvár (Endrédi and Horváth 1976). We cut 20 × 20 × 20 cm sods out of the plant stand and placed them in a photostat. In the photostat the daily rhythm of temperature was 18–28 °C, the relative humidity of air ranged between 45 and 80 per cent, and the carbon dioxide content corresponded to the natural concentration (0.033 per cent). The amount of water supplied to the plants in the form of distilled water every day was nearly the same throughout the experiment.
Three kinds of light treatment were applied. The plant stands were illuminated with blue, red and as a control compound "white" light by means of 40 W Tungsram type F-blue-. F-red- and F33 fluorescent lamps for 12 hours a day. The energy content of the light was the same, about 60 W ∙ m–2, with all three spectral compositions.
At the beginning of the experiment the plant stands were cut back to the ground level. It was then that the species composition and cover of the vegetation were established. Of the samples taken from each subassociation those were placed in the photostat for which the species composition and cover were found to be practically the same, whereby the possibility of comparing them for drymatter production – by the fraction too – obtained with the different light treatments was ensured.
The effect of the spectral composition of light was studied in three experiment series representing the replications, first in 1978, then in 1981 and 1982, under perfectly identical conditions.
The plant stands were raised from 1 April over 120 days each year. The first samples were taken after 60 days, while the second occasion of sampling was at the end of the experiment period. The plants were cut on each occasion from half of the area of the culture pot (2 dm2). In the material of the samples the living and dead plant parts were separated; the living material was sorted into fractions of dominant or prevalent species and of other species, respectively. The living parts represent the net aboveground phytobiomass production, while the living and dead plant parts together the net phytomass production; both are given in dry weight (Précsényi 1975). For the determination of the amount of drymatter the material of the samples was dried at 80 °C to body-balance.
For lack of the necessary equipment there was no possibility to determine the energy content of the plant parts for the estimation of the energy utilization or ecological efficiency, therefore data of the relevant literature were taken into consideration (Golley 1961, Lieth 1968, Moir 1969, Précsényi 1975, etc.). On this basis 1 g drymatter of plant was regarded as equivalent 17580 J energy.
The chlorophyll concentration was determined in those species of each subassociation which had the largest share from the aboveground phytobiomass. They were (Soó 1968, 1970, 1973, 1980, Soó and Kárpáti 1968):
– Salvio-Festucetum rupicolae pannonicum festucetosum rupicolae: Festuca rupicola Heuff.; – S.-F. r. p. stipetosum capillatae: Stipa capillata L. and Salvia nemorosa L.;
– S.-F. r. p. andropogonetosum: Bothriochloa ischaemum (L.) Keng.;
– S.-F. r. p. poëtosum capillatae: Poa angustifolia L. and Achilles collina Becker in Rchb.
The photosynthetic pigments were extracted with acetone – with several replications –from the fresh Alant material at the time of sampling, then washed over into petroleum ether.
The absorption of the pigment extract was measured by a Spekol spectrophotometer at 647 and 664 nm wave-lengths, and the amount of chlorophyll (chlorophyll a + b) determined after Ziegler and Egle (1965). In the course of processing the data the average values of repeated measurings were taken into account. For the sake of a more reliable comparison – knowing the drymatter concentrations – we converted the measuring data obtained for unit weight of fresh material to dry weight figures too.
The variance analysis and the correlation examinations employed in evaluating the experiment results were based on the work of Sváb (1973).
In the paper we reckon with the mean values of the results of the three experiment series, since no significant differences were found between the data obtained in the different years.
Results
Aboveground drymatter production and energy utilization
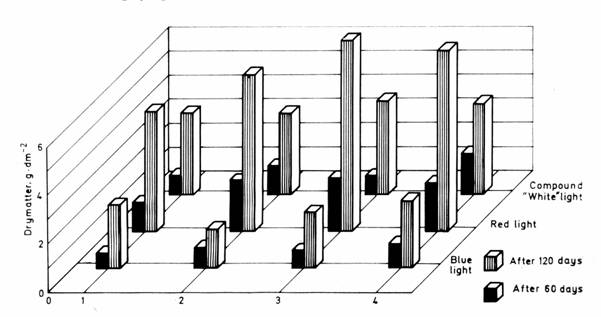
The quantitative data of aboveground phytomass production, separated to fractions, at the time of sampling are summarized by stand and light treatment in Table 1 and represented in Fig. 1.
The largest amount of drymatter was obtained with plant stands raised in red light (the average weight of phytomass was 1.8782 g · dm–2 on the first and 6.7032 g ∙ dm–2 on the second occasion of sampling), the smallest one with those grown in blue light (mean values: 0.8060 g · dm–2 and 2.3466 g ∙ dm–2 respectively). The amount of drymatter produced in compound "white" light was closer to that obtained in blue light (1.1123 g · dm–2 and 3.6132 g ∙ dm–2, respectively).
Fig. 1. Aboveground drymatter production.
1. Salvio-Festucetum rupicolae pannonicum festucetosum rupicolae, 2. S.-F. r. p. stipetosum capillatae, 3. S.-F. r. p. andropogonetosum, 4. S.-F. r. p. poëtosum angustifoliae
The share of the phytobiomass of each species from the total aboveground phytobiomass is shown-according to the different light treatments-in Fig. 2.
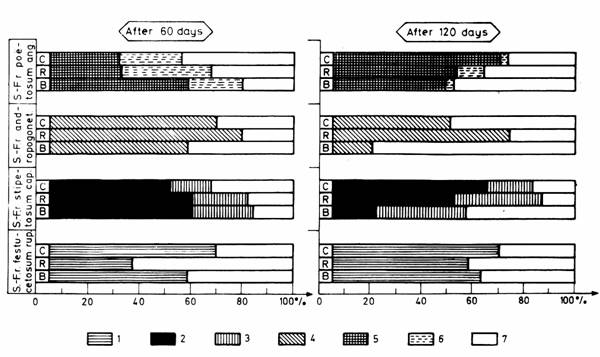
Fig. 2. Share of the phytobiomass of species in the aboveground phytobiomass production.
1. Festuca rupicola, 2. Stipa capillata, 3. Salvia nemorosa, 4. Bothriochloa ischaemum,
5. Poa angustifolia, 6. Achillea collina, 7. Phytobiomass of other species;
B: blue light, R: red light, C: compound "white" light

Table2

Out of the species occurring in the highest proportion Festuca rupicola and Stipa capillata had the largest share in the drymatter of living plant parts when grown in compound "white" light, Salvia nemorosa and Poa angustifolia when raised in blue light, and Achillea collina and Botriochloa ischaemum when treated with red light.
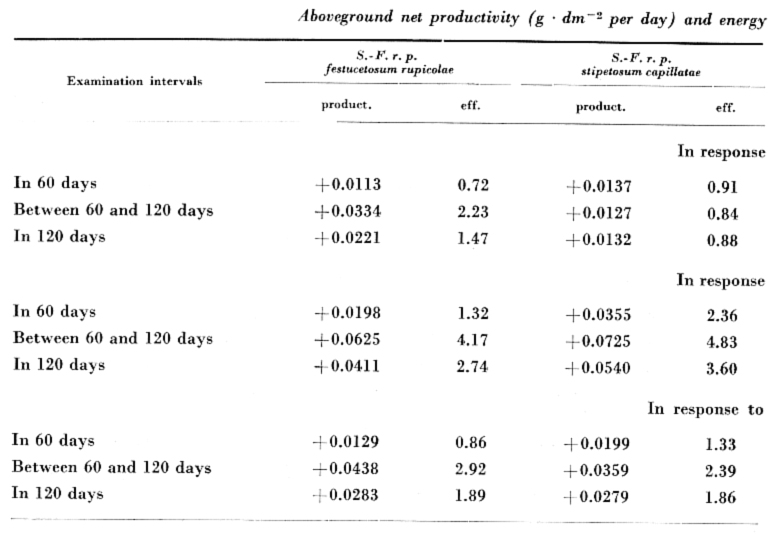
For net productivity on the basis of aboveground phytomass, and energy utilization relative to the total radiation (efficiency) detailed data are given in Table 2. Considering the full period of the experiment, the highest average productivity was obtained in red light (daily value: +0.0559 g · dm–2), the lowest as an effect of blue light (daily average +0.0196 g ∙ dm–2), while in the case of compound "white" light the daily mean value of productivity was +0.0310 g · dm–2. The proportions of energy utilization are similar. The values of efficiency: 3.72 per cent in red light, 1.30 per cent in blue light, and 2.01 per cent as the effect of compound "white" light used.
Chlorophyll contents in the dominant species
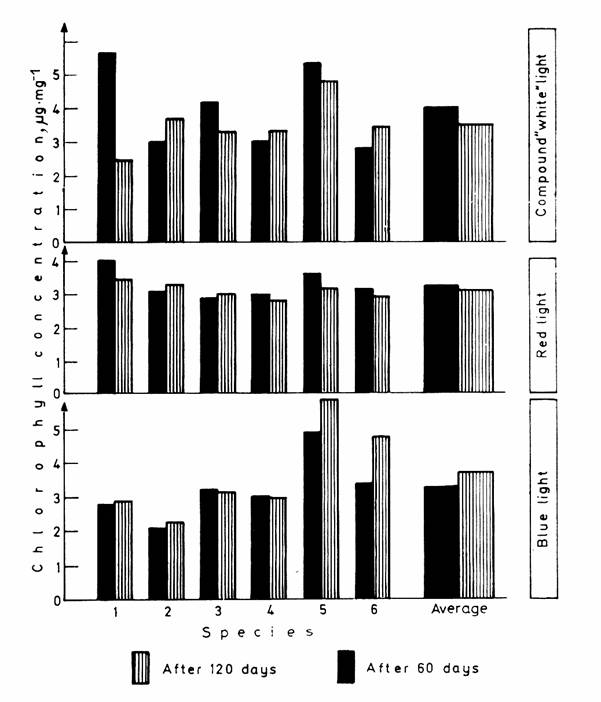
Fig. 3. Chlorophyll concentrations characteristic of the dominant species
in the fresh plant material.
1. Salvia nemorosa, 2. Achillea collina, 3. Festuca rupicola, 4. Poa angustifolia,
5. Stipa capillata, 6. Bothriochloa ischaemum
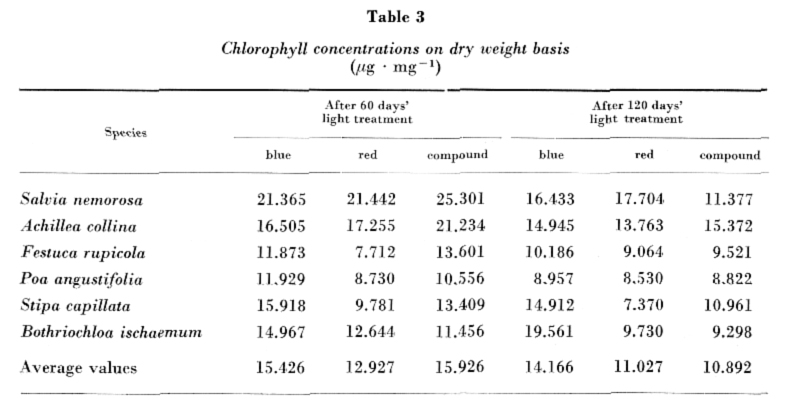
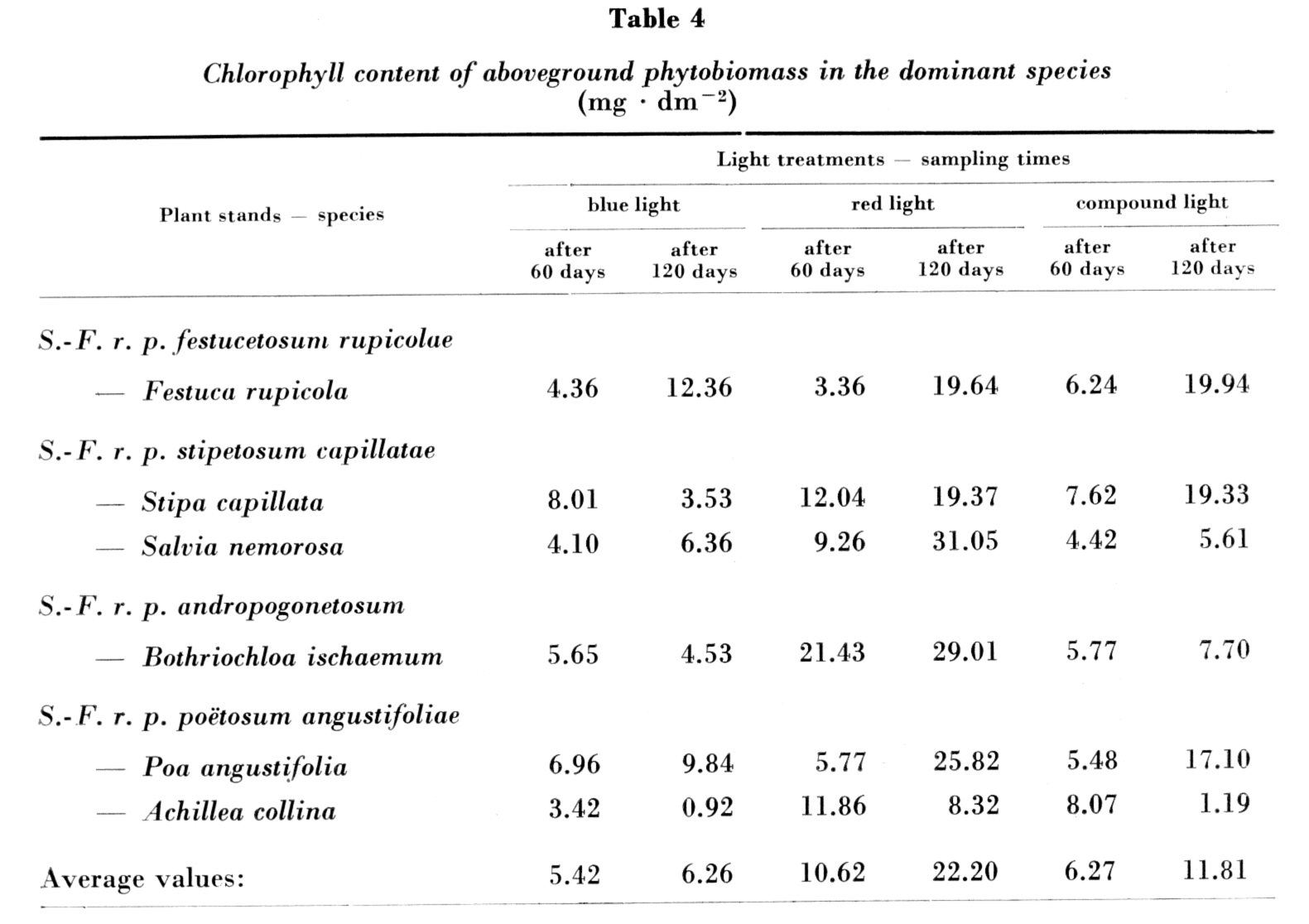
The chlorophyll a + b concentrations per unit weight of the fresh plant material in the six species examined are given in Fig. 3. In general, the differences were not great between the light treatments, the mean values were nearly identical: 3.240 μg ∙ mg–1 with blue, 3.268 μg ∙ mg–1 with red and 3.993 μg ∙ mg–1 with compound "white" light after 60 days; 3.672 , μg ∙ mg–1 with blue, 3.129 μg ∙ mg–1 with red and 3.480 μg ∙ mg–1 with compound "white" light at the end of the experiment.
Chlorophyll concentrations calculated for dry weight are contained in Table 3; the lowest, highest and average values obtained with the light treatments are shown in Fig. 4. As for the mean values of the chlorophyll contents of species on dry weight basis, differences between the light treatments are not considerable either. At the end of the experiment the chlorophyll concentrations were almost in every case lower than those determined after 60 days. The decrease was the greatest in the case of the compound "white" light, while in plant species raised in blue and red light it was negligible.
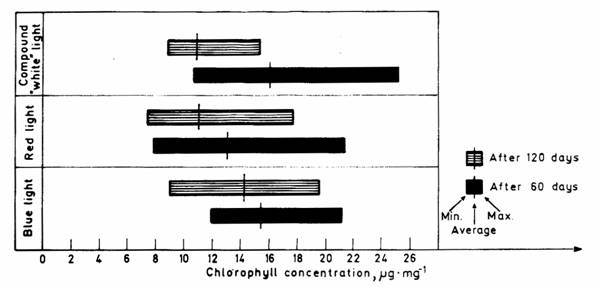
Fig. 4. Lowest, highest and average chlorophyll concentrations
of the six dominant plant species on dry weight basis
The chlorophyll contents of the phytobiomass of species having a decisive or relatively large share in the aboveground drymatter production, as calculated from concentrations characteristic of the dry weight are summed up in Table 4. In Fig. 5 the amounts of phytobiomass and chlorophyll per unit area are shown.
Fig. 5. Aboveground phytobiomass production in the dominant species
and chlorophyll content of the phytobiomass.
1. Festuca rupicola, 2. Stipa capillata, 3. Salvia nemorosa, 4. Bothriochloa ischaemum, 5. Poa angustifolia, 6. Achilles collina; a: blue light, b: red light, c: compound "white" light; A: after 60 days, B: after 120 days
Discussion
The amounts of drymatter produced in blue, red and compound "white" light were compared by variance analysis. The results are given in Table 5.
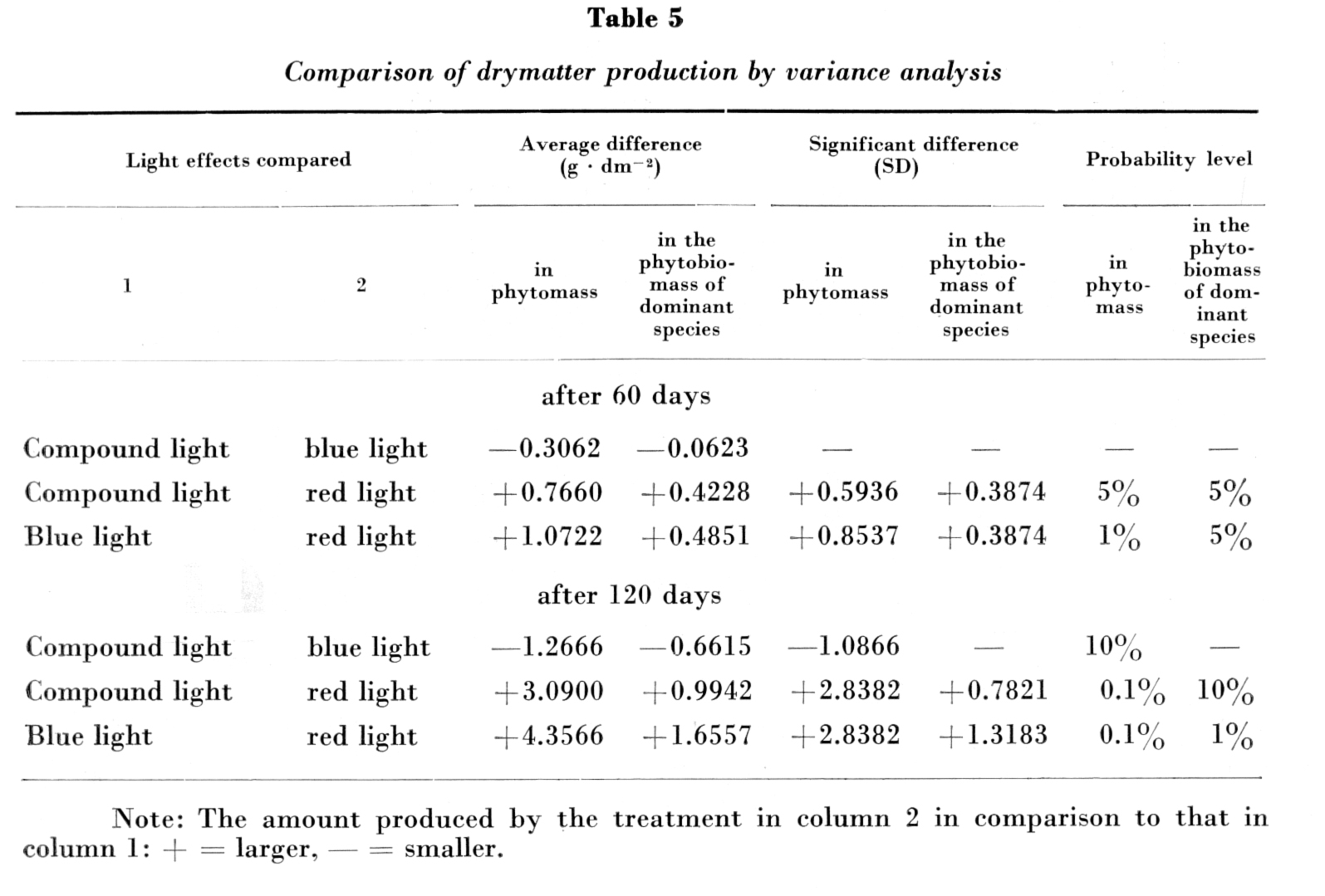
Under the influence of lights of different spectral composition significant differences were found – after 60 and 120 days alike – to occur both in the amount of the phytomass and between the phytobiomass quantities of the dominant species. The greatest difference in average net production was pointed out between plant stands grown in blue and red light, as well as between those raised in compound "white" and red light, especially at the end of the experiment. The amounts of phytomass produced in compound "white" and blue light, respectively, showed essential differences only on the second occasion of sampling, and even then at a low (10 per cent) level of probability. Between the dominant species no significant differences in the amount of phytobiomass were found here. In blue light smaller amounts of phytomass and phytobiomass were produced than in compound light.
The trend of phytomass production observed in our experiment as a response to various spectral compositions of light tan be compared to other authors' results only with restrictions. The comparison is made difficult by the mostly essential differences in the conditions of the experiments, first of all in the period of examination and the energy content of light, but difficulties are also caused by the fact that the drymatter data ere not generally given in terms of unit area. In spite of this the phytomass production obtained in the different light treatments in our experiment followed much the same order as established by the authors listed in the introduction. For example, Hoover (1937), Gabrielseiv (1948), Vince and Stoughton (1957) and Kleschnin(1960) found that with a radiation of the same energy the intensity of photosynthesis in the leaves of many dicotyledonous and monocotyledonous plants was substantially greater in the orange and red wave ranges than in the blueviolet one. Kwack and Dunn (1961) observed with Pisum sativum, Stevenson and Dunn (1965) with Lycopersicon esculentum and Szász, Horváth and Sz. Barsi (1968) with Glycine soja that at the same energy level and light intensity, respectively, a larger amount of drymatter was produced in red than in blue light. In the experiments of Warrington and Mitchell (1976) – including the species of Trifolium, Glycine, Sorgum and Lolium – an increase in the red wave range of the compound "white" light resulted in a similar increase in the dry weight of plants, while with the growing proportion of the blue colour the amount of drymatter decreased.
As to such results as differing from our findings (e.g. Voskreseivskaya and Grishina 1959, Kleschnin 1960, Horváth and V. Fehér 1963, Koltay and Horváth 1967) it is worth being mentioned that the spectral composition of light acts on the primary production in close correlation with the energy level. At different energy level the spectral effect may be highly varied – often even the opposite; and it is also important that the drymatter production in the different species is not uniformly influenced by the spectral composition of light (Horváth 1965, Koltay and Horváth 1967). The latter has been proved bt' the results of our experiment too. The species composing the plant stands gave differentiated responses to the spectral composition, as clearly seen from the fact that their share in the phytobiomass production generally varied with the light treatment, and – with some of them (with Poa angustifolia and Stipa capillata in particular) – even depended on the age of the plants.
We found that the energy of the red light was much better utilized than that of the blue light. It can probably be explained by the fact that at the same energy level – a condition in our experiment too – the number of photons of the red light is about one and a half times higher than that of the blue light, and the in vivo light absorption of the leaves of plants is also greater in the red spectral range than in the blue one (Nuernbergk 1961), so that the chlorophyll molecules can absorb an essentially larger number of photons in red light. Namely, the amount of net drymatter produced depends first of all on the number of photons fixed by' the plants, as proved e.g. by the experiments of Horváth et al. (1973), who obtained the same extent of drymatter accumulation in Sinapis alba plants when the energy of the blue light was 1.79 times that of the red light. This kind of light treatment ensured that the in vivo light absorption of the leaves was almost uniform concerning the number of photons. The light energy fixed was then higher in blue than in red light, but the energy utilization was essentially lower.
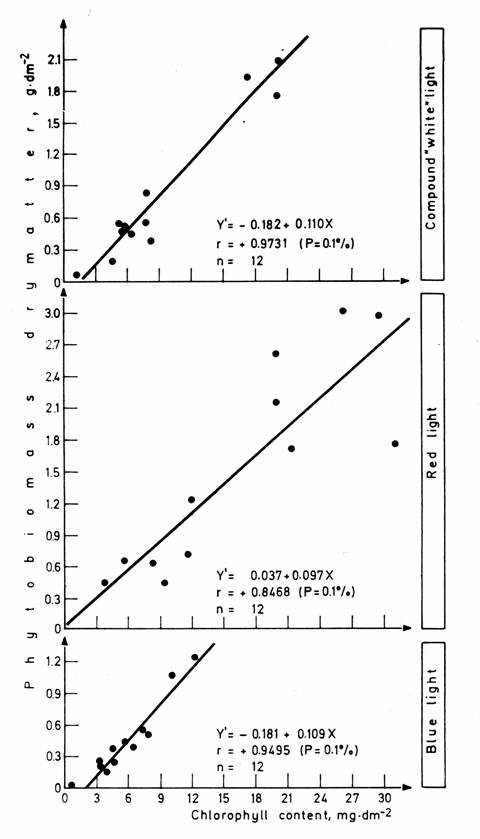
Fig. 6. Relationship between the aboveground phytobiomass production
and the chlorophyll content of the phytobiomass in the dominant species
The six plant species that had the greatest share in the phytobiomass production showed no significant differences in response to various light treatments, in respect of either the average values of chlorophyll content in the fresh plant material, or the average chlorophyll concentrations on dry weight basis. This was most conspicuous in the red light treatment in which the largest amount of drymatter was produced; at the same time the average chlorophyll concentration in the fresh plant material was the lowest under the influence of red light: in the first half of the experiment its value on dry weight basis was lower than in the other treatments, and even at the end hardly exceeded the minimum value measured with compound light. This again suggests that the productivity of the vegetation greatly depends on the number of photons absorbed by the chlorophyll molecules.
We tried to find correlation between the phytobiomass productions per unit area in the dominant species and their chlorophyll contents with regression analysis. The results are shown in Fig. 6. Accordingly there is a very close linear regression correlation between the phytobiomass quantities and the chlorophyll contents. The values of the correlation coefficient are: r = +0.9731 with the compound "white", r = +0.9495 with the blue and r = +0.8468 with the red light. The probability level of significance is in each case 0.1 %. A similar close positive correlation was pointed out between the drymatter production and the chlorophyll content e.g. by Brougham (1960), Whittaker and Garfine(1962), Medina and Lieth (1963, 1964), Ovington and Lawrence (1967) and Endrédi and Horváth (1977) and between the amount of drymatter and that of chlorophyll-a by Rigau and Berbel (1972) and Rigau (1977).
REFERENCES
Brougham, R. W. (1960): The relationship between the critical leaf area, total chlorophyll content, and maximum growth-rate of some pasture and crop plants. Ann. Bot. 24: 363–474.
Endrédi, L. (1981): Összefüggés löszpusztatéri növényzet fitobiomassza-produkciója és a klorofilltartalom között, a fény eltérő színképi összetételének hatására (Relationship between the phytobiomass production and chlorophyll content of a loess-sward vegetation in response to lights of different spectral composition). (Abstract of lecture.) Bot. Köztem. 68: 298–299.
Endrédi, L. (1984): Mezőföldi löszpusztaréti növényállományok talajszint feletti szervesanyagprodukciója és energiahasznosítása szabadföldi körülmények között (Aboveground organic matter production and energy utilization in loess-sward plant stands of "Mezőföld", Transdanubia). A Kaposvári Tanítóképző Főiskola Tudományos Közleményei 6: 89–107.
Endrédi, L., Horváth, I. (1975): Fényszekrény (fotostat) növényökológiai és növényélettani vizsgálatokhoz (Photostat for plant ecology and plant physiology studies). Bot. Köztem. 62: 299–303.
Endrédi, L., Horváth, I. (1976): Organic matter production and photosynthetic energy utilization of plant association in loess grassland. Acta Bot. Acad. Sci. Hung. 22: 39–49.
Endrédi, L., Horváth, I. (1977): Correlation between above-ground phytomass production and the chlorophyll content in the vegetation of a "löszpusztarét", in field experimentation and in conditioned situations. Acta Bot. Acad. Sci. Hung. 23: 63–75.
Endrédi, L., Horváth, I. (1978): A fény színképi összetételének hatása a löszpusztaréti növényzet talajszint feletti produkciójára (Influence of the spectral composition of light on the aboveground production of a loess-sward vegetation). A XIII. Biológiai Vándorgyűlés Előadásainak Ismertetése. (Abstract of a lecture.) Magyar Biológiai Társaság, Budapest, 90.
Fekete, G. (1972): A növénytársulás fiziognómiai struktúrája, a fény és a víz mint produkcióökológiai tényezők (Physiognomical structure of plant association, light and water as factors of production ecology). MTA Biol. Oszt. Közt. 15: 137–158.
Gabrielsen, E. K. (1948): Influence of light of different wave-length on photosynthesis in foliage leaves. Physiol. Plant. 1: 113–123.
Golley, F. B. (1961): Energy values of ecological materials. Ecology 42: 581–583.
Hauschild, A. H. W., Nelson, C. D., Krotkov, G. (1961): Effect of light quality on the products of photosynthesis in Chlorella vulgaris. Plant Physiol. 36: Suppl. XXXVI.
Hauschild, A. H. W., Nelson, C. D., Krotkov, G. (1964): Concurrent changes in the products and the rate of photosynthesis in Chlorella vulgaris in the presence of blue light. Naturwiss. 51: 274–275.
Hoover, W. H. (1937): The dependence of tarbon dioxide assimilation in a higher plant on wavelength of radiation. Smithsonian Misc. Coll. 95 No. 21.
Horváth, I. (1975): A fény színképi összetételének növényökológiai szerepe (Role of the spectral composition of light in plant ecology). Thesis, Budapest.
Horváth, I., V. Fehér, I. (1963): Különböző színképi összetételű fényben nevelt növények fotoszintetikus pigmentkomplexének fényabszorpciója (Light absorption by the photosynthetic pigment complexes of plants raised in lights of different spectral composition). Agrártud. Egyetem Mezőgaxdaságtud. Kar Közl. 397–404.
Horváth, L, Szalay, L., Szász, K., Raafat, A. (1973): The effect on the spectral composition of light on the metabolism. Utilization of light and the chlorophylls of Sinapis alóa. Acta Biochim. et Biophys. Acad. Sci. Hung. 8: 161–169.
Horváth, L, Mihalik, E., Takács, E. (1978): Nátriumlámpa hatása a növények szárazanyag produkciójára (Effect of sodium discharge lamp on drymatter production in plants). Bot. Közlem. 65: 115–121.
Horváth, L, Mihalik, E. (1980): Kombinált megvilágítás hatása a növények szárazanyag produkciójára (Effect of combined illumination on drymatter production in plants). Bot. Közlem. 67: 72–82.
Horváth, L, Mihalik, E., Takács, E. (1980): Effect of light on root production. In: Sen, D. N. (ed.): Environment and Root Behavior. Geobios Internat., Jodhpur, India, 231-255. Kadman-Zahavi, A., Alvarez-Vega, E. (1968): The Use of Coloured Light for Agriculture. The National and University Institute of Agriculture, Bet Dagan, Israel.
Kleschnin, A. F. (1960): Die Pflanze und das Licht. Akademie Verlag, Berlin.
Koltay, A., Horváth, I. (1967): A fény színképi összetételének hatása a szárazanyag-produkcióra és lokalizációjára (Effect of the spectral composition of light on the drymatter production and its localization). Agrártud. Egyetem Mezőgaxdaságtud. Kar Közl. 141–151.
Kowallik, W. (1962): Über die Wirkung des blauen und roten Spektralbereiches auf die Zusammensetzung und Zellteilung synchronisierter Chlorellen. Planta 58: 337-365.
Kwack, B. H., Dunn, S. (1961): Effects of light quality on plant maturity. I. Duration of growth, nutrient supply, and photoperiod. Lloydia 24: 75–80.
Lieth, H. (1968): The measurement of calorific values of biological material and the determination of ecological efficiency. UNESCO Copenhagen Symposium. 233–242.
Medina, E., Lieth, H. (1963): Contenido de clorofila de algunas associaciones vegetales de Europa Central y su relación con la productividad. Qualitas Plant. Mat. Veget. 9: 217–229.
Medina, E., Lieth, H. (1964): Die Beziehungen zwischen Chlorophyllgehalt, assimilierender Fläche und Trockensubstanzproduktion in einigen Pflanzengemeinschaften. Beitr. Biol. Pflanzen 40: 451–494.
Moir, W. H. (1969): Energy fixation and the role of primary producers in energy flux of grassland ecosystems. In: Grassland Ecosystem, 125–147.
Nuernbergk, E. L. (1961): Kunstlicht und Pflanzenkultur. BLV Verlags Gesellschaft, München–Bonn–Wien.
Ovington, J. D., Lawrence, D. B. (1967): Comparative chlorophyll and energy studies of prairie, savanna, oakwood, and maize field ecosystems. Ecology 48: 515–524.
Pirson, A., Kowallik, W. (1964): Spectral responses to light by unicellular plants. Photochem. Photobiol. 3: 489–497.
Précsényi, I. (1975): Szikespusztai rét növényzetének produktivitása (Productivity of an Alkali Grassland). Akadémiai Kiadó, Budapest.
Rigau, M. C. (1977): Relationship between production characteristics and chlorophyll a content in Hedysarum coronarium L., Dactylis glomerata L., and Festuca elatior ssp. arundinacea (Schreb.) Hack. Photosynthetica 11: 88–89.
Rigau, M. C., Berbel, M. (1972): Correlaciones de los pigmentos foliares con el crecimiento y la producción. II. Estudio de las correlaciones existentes entre los pigmentos clorofila a y carotinoides y la producción en Lolium perenne L. Publ. Inst. Biol. 52: 81–106.
Soó, R. (1964, 1968, 1970, 1973, 1980): A magyar flóra és vegetáció rendszertani-növényföldrajzi kézikönyve (Systematic-phytogeographic Manual of the Hungarian Flora and Vegetation). Vols I–III–IV–V–VI. Akadémiai Kiadó, Budapest.
Soó, R., Kárpáti, Z. (1968): Magyar flóra. Harasztok. Virágos növények (Növényhatározó. II (Hungarian Flora. Ferns. Flowering plants. (Plant identification book. II)]. Tankönyvkiadó, Budapest.
Stevenson, E. L., Dunn, S. (1965): Plant growth effect of light quality in sequences and in mixtures of light. Advancing Frontiers of Plant Sciences 10: 177–190.
Sváb, J. (1973): Biometriai módszerek a kutatásban (Biometrical Methods in Research). Mezőgazdasági Kiadó, Budapest.
Szász, K. (1967): A fény színképi összetételének hatása a növényi szervesanyag produkcióra (Influence of the spectral composition of light on organic matter production in plants). Thesis, Budapest.
Szász, K., Horváth, L, Sz. Barsi, E. (1968): Differential effect of red and blue light on the accumulation of carbohydrates and nitrogen compounds. Acta Biol. Szeged 14: 35–39.
Tregunna, S. B., Krotkov, G., Nelson, C. D. (1962): Effect of white, red and blue light on the nature of the products of photosynthesis in tobacco leaves. Can. J. Bot. 40: 317–326.
Veen, R, van der, Meijer, G. (1959): Light and Plant Growth. Macmillan Co., New York.
Vince, D., Stoughton, R. H. (1957): Artifical light in plant experimental work. In: Hudson, J. P. (ed.): Control of the Plant Environment. London.
Voskresenskaya, N. P. (1965): Photosynthesis and Spectral Composition of Light. Nauka, Moskwa.
Voskresenskaya, N. P. (1972): Blue light and carbon metabolism. Ann. Rev. Plant Physiol. 23: 219–234.
Voskresenskaya, N. P., Grishina, R. H. (1959): O dejstvü intersivnosti iz spektralnovo sostava radiacii na obmen veschestv i urozanj. Dokl. AN SSSR 124: 469–472.
Warrington, I. J., Mitchell, K. J. (1976): The influence of blue and red biased spectra on the growth and development of plants. Agricult. Met. 16: 247–262.
Whittaker, R. H., Garfine, V. (1962): Leaf characteristics and chlorophyll in relation to exposure and production in Rhododendron maximum. Ecology 43: 120–125.
Ziegler, R., Egle, K. (1965): Zur quantitativen Analyse der Chloroplastenpigmente. I. Kritische Uberprüfung der spektralphotometrischen Chlorophyll-Bestimmung. Beitr. Biol. Pflanzen 41: 11–37.